

1. 서 론
흔히 배스라 불리는 큰입배스(큰입우럭, 민물농어, Largemouth Bass)는 Micropterus salmoides이 학명인 외래어종으로 우리나라에 1970년대에 도입된 것으로 알려져 있다. 큰입배스는 부족한 식량자원을 확보하고 자원을 조성할 목적으로 도입하였는데, 철원에 위치한 토쿄지와 팔당호에서의 방류를 시작으로 현재는 하천 및 호수의 민물생태계에서 빠르게 퍼져 나가고 있다(Hong and Son, 2003; Kang and Kim, 2016; Ko et al., 2008; Park et al., 2019; Yoon et al., 2018). 큰입배스는 대표적인 육식어종으로 어류, 양서류, 갑각류 등 다양한 먹이를 섭식하며, 환경의 변화에 따른 적응력과 증식력이 강한 것으로 알려져 있다. 큰입배스와 함께 대표적인 유해 외래어종으로 분류되는 블루길(파랑볼우럭, Bluegill) 역시 우리나라 전역에서 서식하는 것으로 보고된 바 있다(Yoon et al., 2018).
큰입배스와 블루길의 유입은 우리나라 민물생태계의 생물다양성 감소와 생태계 교란을 야기하는 주요 요인으로 여겨지며(Park et al., 2019; Byeon, 2018), 큰입배스의 도입으로 인해 어류의 종 다양성이 감소한 사례들은 전 세계적으로 보고되고 있다. Maezono and Miyashita (2003)은 용수조절지에서 블루길과 큰입배스의 유무에 따른 수생군집의 변화를 살펴보았는데, 블루길과 큰입배스가 존재하는 경우 피라미, 새우, 잠자리의 유충과 같은 토착생물의 개체수가 현저하게 낮은 것을 확인하였다. 남아프리카에 위치한 호수에서는 큰입배스의 유입으로 인해 가재와 같은 토착생물의 멸종이 보고된 바 있으며(Hickley et al., 1994), 멕시코에서는 스포츠 어업을 위해 이식한 큰입배스로 인해 토착어종의 수가 급감하여 주민의 어업생산율이 감소한 사례 또한 확인된다(Ciruna et al., 2004). 미국 내 저수지에서도 큰입배스의 유입으로 인해 토착생물인 피라미와 토착어종의 개체수가 급감한 사례를 찾을 수 있으며, 캐나다에서도 큰입배스의 유입으로 인해 피라미와 잉어과의 토착 어종이 현저히 감소한 사례가 보고된다(Findlay et al., 2000; Broadway and Moyle, 1978; Kerr and Grant, 2000).
큰입배스로 인한 종 다양성 감소는 민물생태계의 교란을 야기하며, 우리나라 환경부에서는 큰입배스를 생태계 교란생물로 지정하여 법적으로 관리하고 있다. 우리나라에서는 인공산란장을 이용한 알 제거, 작살을 이용한 포획, 천적을 이용하는 방법, 수매 등의 방법을 통해 외래어종 퇴치에 노력을 기울이고 있으나, 외래어종의 개체수와 분포도를 고려할 때 포획에 의한 퇴치방법은 그 실효성에서 의문을 가지게 된다. 뿐만 아니라, 외래어종이 생존을 위해 토종어류를 먹이로 삼는다는 이유 등으로 작살을 이용하여 포획하고 이를 매장하는 방법은 생물의 생명존엄성을 무시하는 대책이라는 비판 역시 존재한다. 해외에서는 외래어종의 퇴치를 위해 특정 종만을 대상으로 효과를 발휘한다는 약물의 적용 등 매우 적극적인 퇴치방안을 시행하였으나 효과를 거두지 못하고 있다. 일본 역시 수생태계의 교란에 큰 관심을 가지며 큰입배스의 퇴치를 위해 노력을 기울이고 있다(e.g., Katano and Sakano, 2010; Maezono and Miyashita, 2003). 그러나 큰입배스의 포획으로 인해 블루길이 증가하였으며, 토착어종이 증가하여 수중식물이 감소하는 문제가 발생하는 것을 현장실험 결과로부터 확인하였다(Maezono et al., 2005). 이상에서 살펴본 바에 따르면 우리나라의 현재 상황에 대처하기 위해서는 포획에 의한 퇴치보다 외래어종의 서식특성에 근거하여 확산을 제어하는 것이 더욱 합리적일 것이라 생각해 볼 수 있다.
우리나라에서 외래어종 또는 특정어종의 확산을 제어하기 위한 연구는 여러 차례 시도된 적이 있다. 현재까지 차단망과 같이 직접적으로 접근하는 방법과 음파를 이용한 간접적인 접근방법을 이용한 연구가 수행되었다. Kang and Kim (2017)은 물거품을 이용한 어류 차단 시스템을 개발하여 외래어종인 블루길을 대상으로 어류의 차단 가능성을 평가하였다. 물거품을 이용한 시설물을 설치한 경우 블루길의 차단 효과가 매우 높았으나, 물거품 발생을 종료할 경우 빠른 시간 내 다시 블루길이 이동하는 사실이 확인되었다(Kang and Kim, 2017). 따라서 물거품을 이용한 어류 차단은 반드시 연속적으로 이루어져야 한다는 점을 알 수 있다. 물거품을 이용한 이동경로의 차단이 단기간에는 효과적일 수 있지만 장기간에 걸쳐 어류가 물거품 환경에 적응하게 된다면 어떤 결과가 나타날 것인가에 대한 의문이 생기며 용존산소가 증가할 수밖에 없는 상황에서 오히려 어류를 유인하는 결과가 도출될 수 있다는 점도 예상된다. 북미가 원산지인 큰입배스의 경우 흐름이 약한 정체구역을 중심으로 서식처를 형성하는 것으로 알려져 있으나(Kim et al., 2013), 하천 중류의 특성이 많이 존재하는 우리나라의 환경에 적응한 지금은 여울 등 흐름이 매우 빠른 환경에서도 큰입배스의 서식처를 쉽게 확인할 수 있다. 따라서 물거품이라는 특이한 환경조건에서의 적응 역시 길지 않은 시간 내에 이루어질 것으로 예상된다. Kang et al. (2014)은 소리를 이용한 어류의 차단 효과를 어류 차단구조물을 개발하여 블루길과 큰입배스를 대상으로 차단 가능성을 살펴보았다. 블루길과 큰입배스 모두 소리에 특이할만한 거부반응이 나타나지 않았는데, 이는 우리나라의 서식환경과 관련이 있는 것으로 판단된다. 우리나라의 서식환경은 잦은 하천공사와 인간 활동으로 인해 소음에 어느 정도 적응할 수밖에 없는 특성을 보인다. 특히 하천 구간의 상당부분이 도로와 인접한 우리나라 특성상 차량으로 인한 진동, 항타 등의 공사 활동 소음에 적응하지 않고는 매우 국한된 서식처만 확보하게 될 것이다. 따라서 음파와 물거품을 이용한 접근법이 효율적인 확산 제어대책으로 생각되지는 않는다.
이에 본 연구에서는 효율적인 외래어종의 확산 제어대책을 수립하기 위한 큰입배스의 서식처 형성 특성을 살펴보고자 하였다. 우리나라에서 수행된 큰입배스에 관한 연구로는 유속에 따른 소상능력을 평가한 연구(Kang and Kim, 2016), 한반도 내 큰입배스의 분포에 관한 연구(Yoon et al., 2018; Byeon, 2018), 흐름 특성에 따른 큰입배스의 식성변화에 관한 연구(Park et al., 2019), 수질 변수가 큰입배스의 서식처 형성에 큰 영향을 미치지 않는다는 연구(Kim et al., 2013) 등이 보고되고 있다. 큰입배스의 서식특성에 관한 연구로는 식성에 대한 연구만이 존재하여 우리나라 민물생태계에 적응한 큰입배스의 기초적인 서식특성에 관한 연구가 절실한 상황이다. 보다 면밀한 관찰을 통해 큰입배스의 서식특성을 검토하기 위해 현장조사를 실시하였으며, 큰입배스의 서식처 형성과 수온이 상관성을 나타낸 현장조사 결과를 바탕으로 실험실 실험을 수행하였다.
2. 연구 방법
2.1 조사지점 및 시기
현장조사는 우리나라 담수 환경의 대표적인 물리적 특성을 살펴볼 수 있도록 여러 지점에서 수행하였다. 현장조사 지점을 크게 구별한다면 정체수역인 저수지권과 흐름이 존재하는 하천권으로 구별할 수 있다. 현장조사를 위해 조사용 보트를 활용하거나 선박장비를 임대하였고, 휴대용 어군탐지기와 수중카메라를 이용하여 어류의 분포특성을 파악하였다. 분포어류의 어종 파악이 불가능한 어군탐지기와 수중카메라의 한계를 극복하기 위해 낚시를 이용한 채집도구를 구축하여 어종을 파악하였다. 낚시를 채집방법으로 선정하기 이전에 족대와 어망을 통한 포획을 고려하였으나, 계곡형 저수지의 깊은 수심에서의 이용이 불가능하다는 점과 서식처 변형을 최소화하기 위해 낚시를 채집방법으로 선정하였다. 늦은 춘계부터 이른 하계까지의 약 9개월간 총 20회의 현장조사를 실시하여 계절별 어류 분포 특성을 살펴보았다. 물이 어는 시기에는 현장조사 대신 간단한 답사의 과정을 통해 서식처 변화양상을 파악하였다. 현장조사 시 어류의 분포, 어종, 서식처의 물리적 특성, 당일의 기상조건, 수심별 수온, 지점별 평균 수온을 조사하였다.
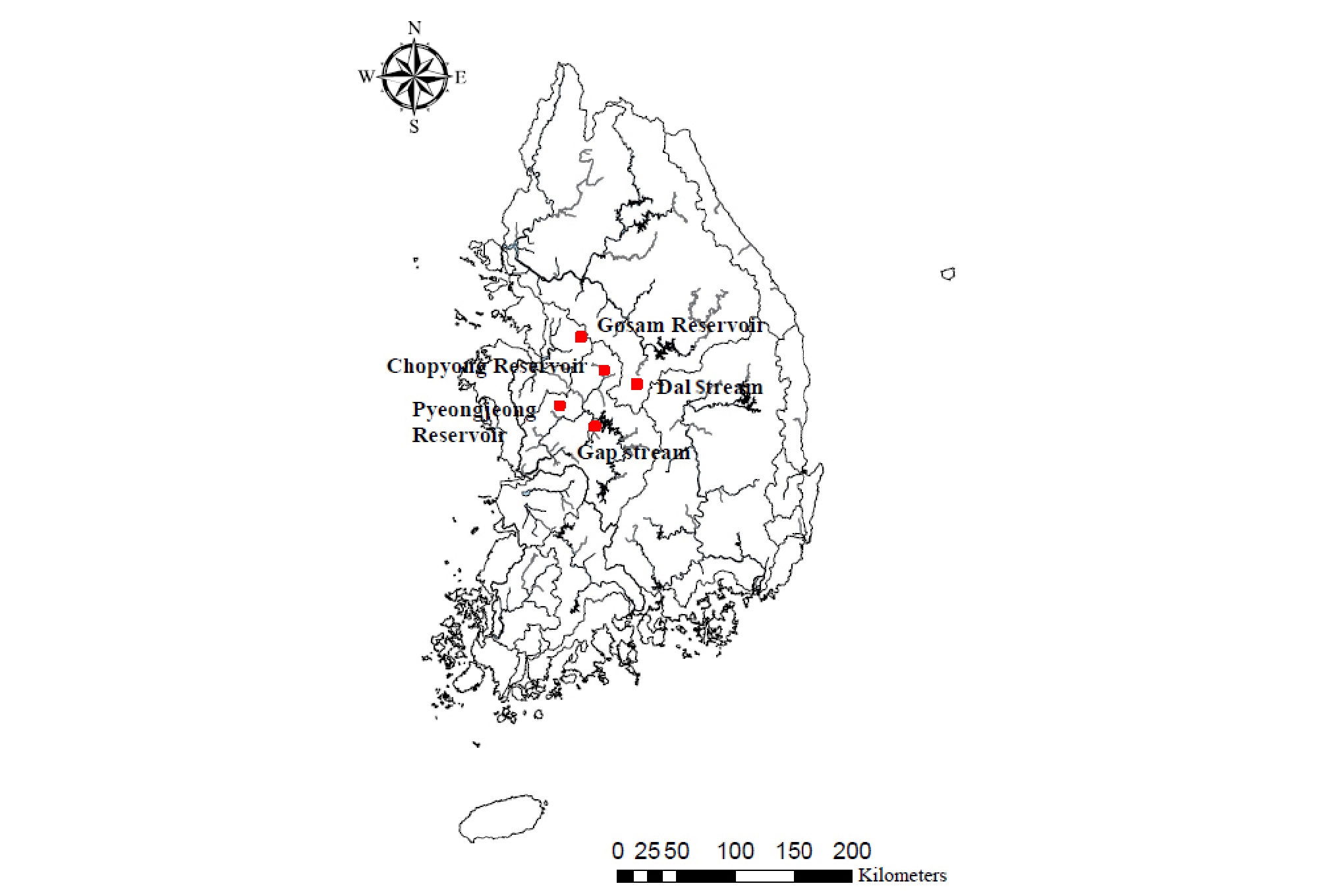
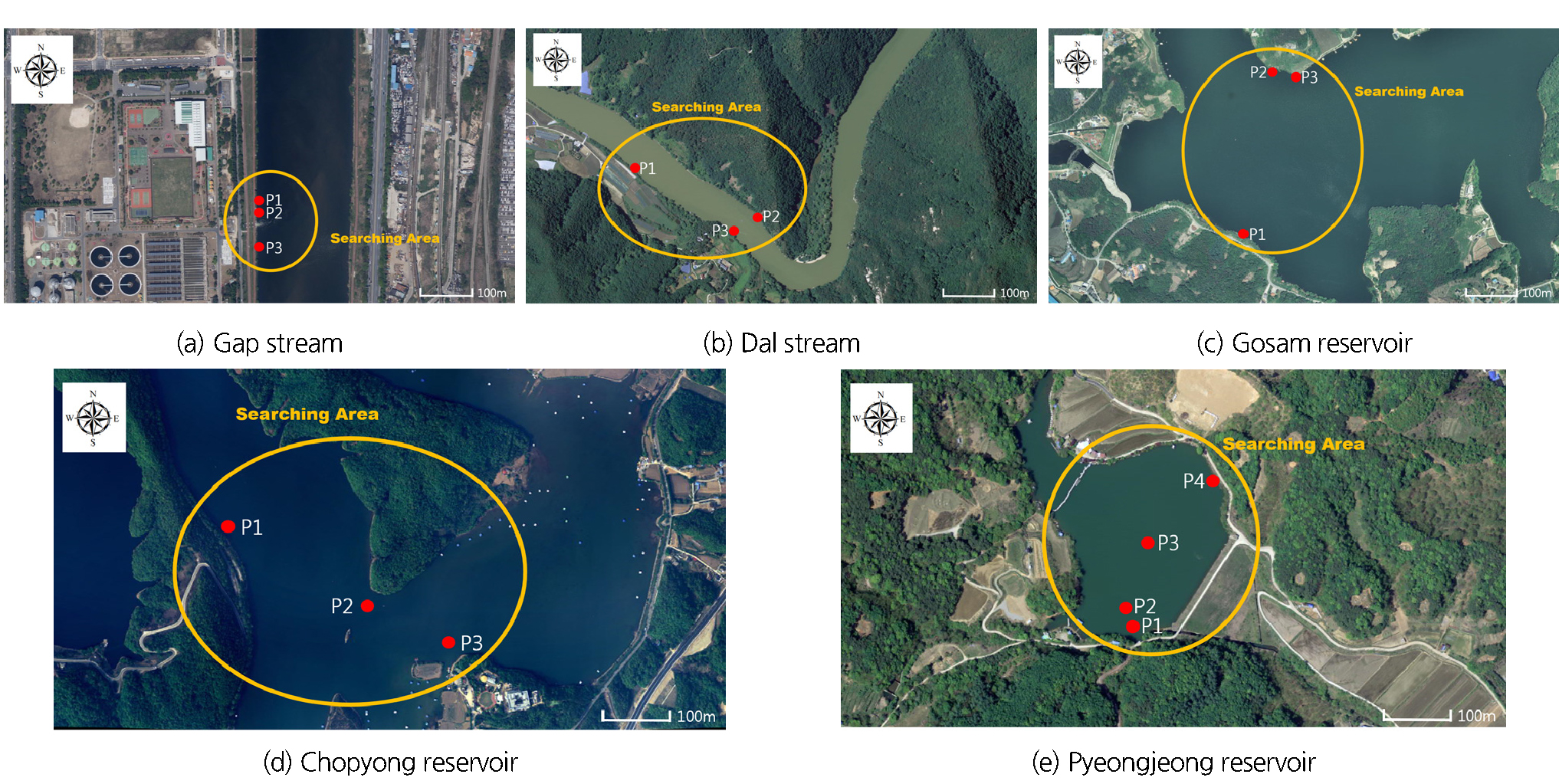
Fig. 1은 현장조사가 수행된 여러 지점을 나타낸다. Figs. 2(a) and 2(b)는 각각 흐름이 존재하는 하천권에서 어류의 분포특성 및 수온을 살펴본 지점을 나타내며, Fig. 2(c) 부터 Fig. 2(d)까지는 정체수역인 저수지에서 수행한 현장조사 지점이 확인된다. Fig. 2(a)는 대전광역시 유성구 원촌동 24-1번지 인근의 갑천 지점을 나타낸 것으로, 도시하천의 특성을 보이는 구간을 선정하였다. 이 지점에는 소화 등의 과정을 거치며 수온이 증가한 하수의 방류가 일어나는 하수처리수의 방류구가 존재한다. 하수 유입부에서 상류방향 및 하류방향으로 각각 약 30 m씩 60 m에 걸친 구간에 대하여 11월에 현장조사를 실시하였다. Fig. 2(b)는 충북 괴산군 청천면 운교리 600번지에 위치한 달천 상류부에서 현장조사를 실시한 지점을 나타내며, 달천에서 수행된 현장조사는 비교적 따뜻한 봄철에 수행되었다. Fig. 2(c)은 경기도 안성시 고삼면의 고삼저수지 내 현장조사가 시행된 지점이며, Fig. 2(d)은 충북 진천군 초평면 화산리에 위치한 초평저수지 내 현장조사가 실시된 지점을 나타낸다. 고삼저수지와 초평저수지에서는 각각 1월 4일과 11월 27일에 현장조사를 실시하였다. Fig. 2(e)은 집중조사를 시행하기 위해 선정된 평정저수지를 나타낸 그림이다. 평정저수지에서는 2015년 11월 20일에서 2016년 6월 21일까지 총 11회의 현장조사가 실시되었다.
2.2 실험실 서식처 구축
실험실 실험을 위해 충남대학교 수공학연구실에서 보유한 길이 12 m, 폭 0.92 m, 높이 0.6 m의 대형수로를 이용하였다. 모래, 자갈, 호박돌을 실험 수조의 아래층부터 순차적으로 설치하여 입도분포가 양호하도록 하상토를 구성하여 자연 상태와 유사한 환경을 실험 수조에서 모사하고자 하였다(Fig. 3(a)). 수로의 좌측 1/3 구간은 수온을 증가시킬 수 있는 가열장치를 모래층과 자갈층 사이의 바닥에 매설하였다. 수로의 우측 1/3 구간에는 수온을 저하시킬 냉각장치를 설치하였으며, 중앙 1/3 구간에는 온도조절장치를 설치하지 않았다(Fig. 3(b)). 각 구간마다 생태환경에서 지배적인 역할을 하는 용존산소량이 본 연구의 진행에 영향을 미치지 않게 충분한 산소공급장치를 설치하였고, 검은 천막을 이용하여 빛을 차단하여 외부의 간섭을 최소화 하고자 하였다. 수조의 윗부분에 광도의 조절이 가능한 조명을 설치하여 태양광선을 대신하였다(Fig. 3(c)).
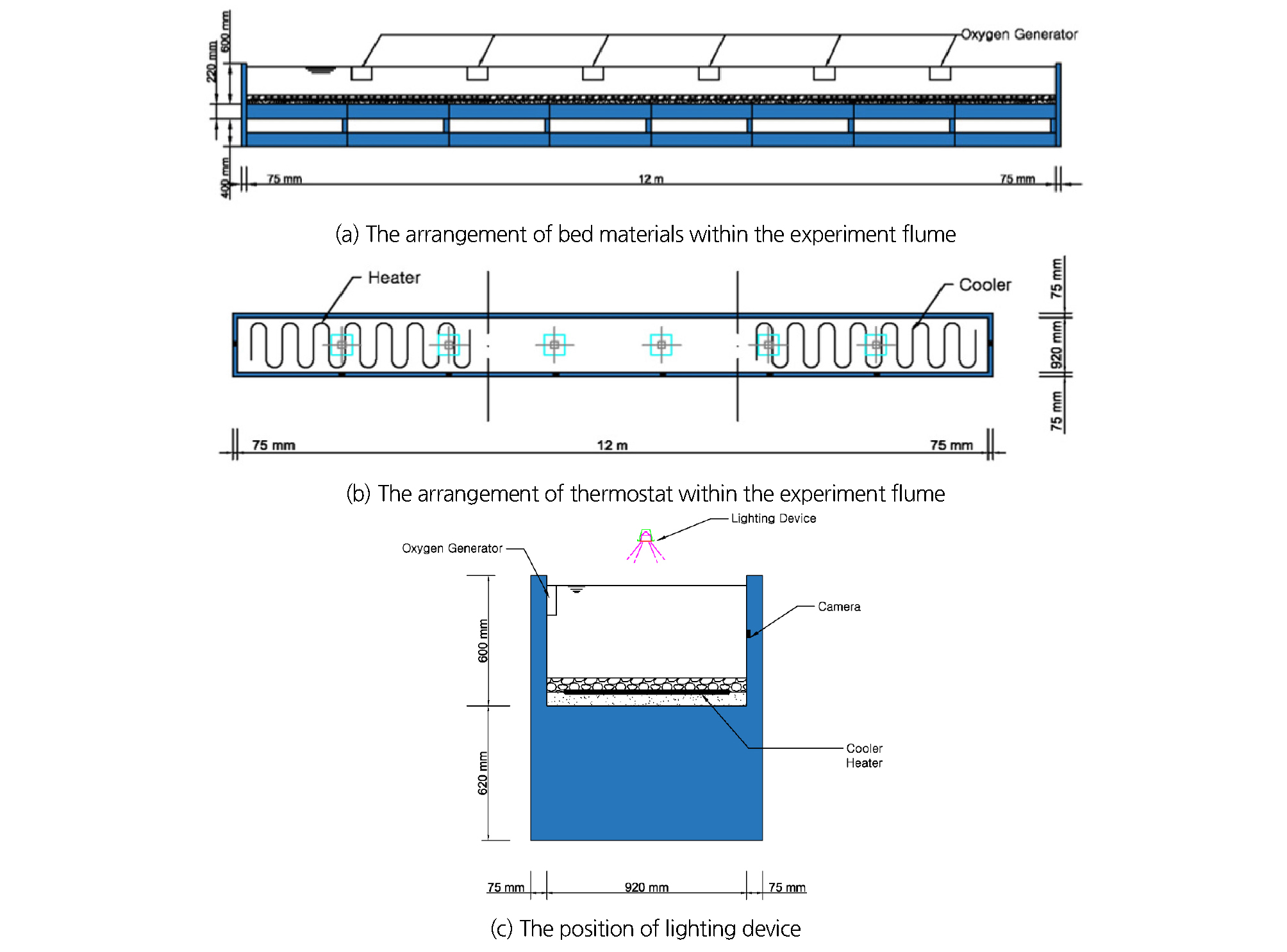
토종어종과 외래어종을 같은 공간에서 사육하여 서식처의 분할과 먹이경쟁, 산란 특성의 변화를 관찰하기 위해서 수로에는 큰입배스, 블루길 등의 외래어종 이외에 토종 육식어종인 붕어도 이식하여 실험을 수행하였다. 외래어종과 토종 육식어종의 채집을 위해서 현장답사를 실시하였으며, 채집된 외래어종 및 토종어종은 환경부의 허가 아래 산소발생기가 설치된 낚시 가방에 넣어 안전하게 실험실 수조에 이식하였다. 이동 과정에서 발생할 수 있는 채집으로 인한 생태영향을 최소화하기 위해 노력하였다. 실험실 서식처에서 외래어종의 생태환경이 수온에만 지배받을 수 있도록 바닥 지형, 채광 등과 같은 조건은 세부구간에서 동일하게 조성하였다. 수조 내에서 수심별로 수온이 달라 생길 수 있는 오차의 영향을 줄이기 위해 구간마다 두 개의 온도계를 각각 다른 수심에 설치한 후, 그 값을 산술평균하여 수온으로 측정하였다. 실험에 적합한 수질 상태를 유지하기 위해 여과기능 장치를 가진 정화장치 5식을 설치하였다.
3. 결과 및 논의
3.1 현장조사
갑천 지점에서 2015년 11월 19일에 수행된 현장조사 결과 하수 방류구를 기준으로 하여 상하류 각각 30 m 지점에서 수온은 약 2℃의 차이를 나타내는 것으로 확인되었다. 방류구를 기준으로 수온이 더 높은 하류 지점에서만 큰입배스가 채집되는 것으로 보아 11월 말 하천에서의 서식처 형성이 고수온 지역을 중심으로 이루어짐을 알 수 있다. 2015년 12월 4일 동일한 지점에서 재조사를 실시하였을 때의 상하류 수온 차이는 2.5℃(상류 11℃, 유입되는 처리수 15℃, 하류 13.5℃)로 나타났으며 1차 조사와 동일하게 하류에서만 채집이 가능하였다.
달천 상류부에서 수행한 현장조사의 시기는 외래어종이 활발한 먹이활동과 산란을 준비하는 행동을 보이는 4월 초순이다. 총 5개 지점에서 정밀조사를 실시한 결과로부터 지점 2를 제외한 4개 지점 모두에서 어류의 분포와 어종이 큰입배스임을 확인할 수 있었다(Fig. 2(b)). 지점 1과 지점 2는 16.0℃로 동일한 수온을 나타내었고 지점 5에서 15℃에 해당하는 최저 수온이 측정되었다. 이를 통해 춘계의 경우 동계와 달리 1℃의 온도 차이가 외래어종의 서식처 형성에 비교적 미미한 영향만을 미치는 것을 알 수 있다. 어류의 분포가 각각 존재, 부재하는 지점 1, 2의 연직 수온 분포를 조사한 결과에 따르면, 수온의 연직 분포가 큰 차이를 보이지 않는 점을 확인할 수 있다. 이를 통해 수온이 15℃ 정도인 춘계의 경우 어류 서식처의 형성 여부가 수온에 큰 영향을 받지 않음을 재확인할 수 있다.
고삼저수지에서는 2016년 1월 4일 겨울철에 현장조사를 수행하였는데, 지점 1부터 3까지의 수심 1 m에서의 수온은 각각 4.2℃, 4.4℃, 5.5℃로 차이를 보였다. 지점 2와 지점 3은 인접하나(Fig. 2(c)), 세 조사지점 중 수온이 1.1℃이상 높은 지점 3에서만 어류가 분포하는 것으로 확인되었다. 표층수온이 4℃ 이하일 때 활동을 거의 정지하는 것으로 알려진 외래어종의 특성을 고려할 때 1.1℃의 수온차가 서식처를 형성하는 과정에 큰 영향을 주는 것으로 예상해 볼 수 있다.
초평저수지에서 2015년 11월 27일에 수행된 현장조사 결과에 따르면, 초평저수지의 수온은 약 10℃로 관측되었다. 저수지 내 수온은 하류로 갈수록 상승하는 것으로 조사되었으며, 지점 1부터 3까지의 수온은 각각 10℃, 9℃, 8℃로 차이가 있는 것으로 나타났다(Fig. 2(d)). 가장 수온이 낮은 지점인 3의 경우에는 어류 활동이 나타나지 않는 대신 지점 1에서는 다수의 어류 분포를 관찰하였으며(Fig. 4(a)), 관찰된 어류의 어종이 큰입배스임을 확인하였다(Fig. 4(b)). 또한 지점 1을 이외 수온이 약 10℃로 측정된 곳에서는 어탐기를 통해 많은 어군 활동을 관찰할 수 있었다.

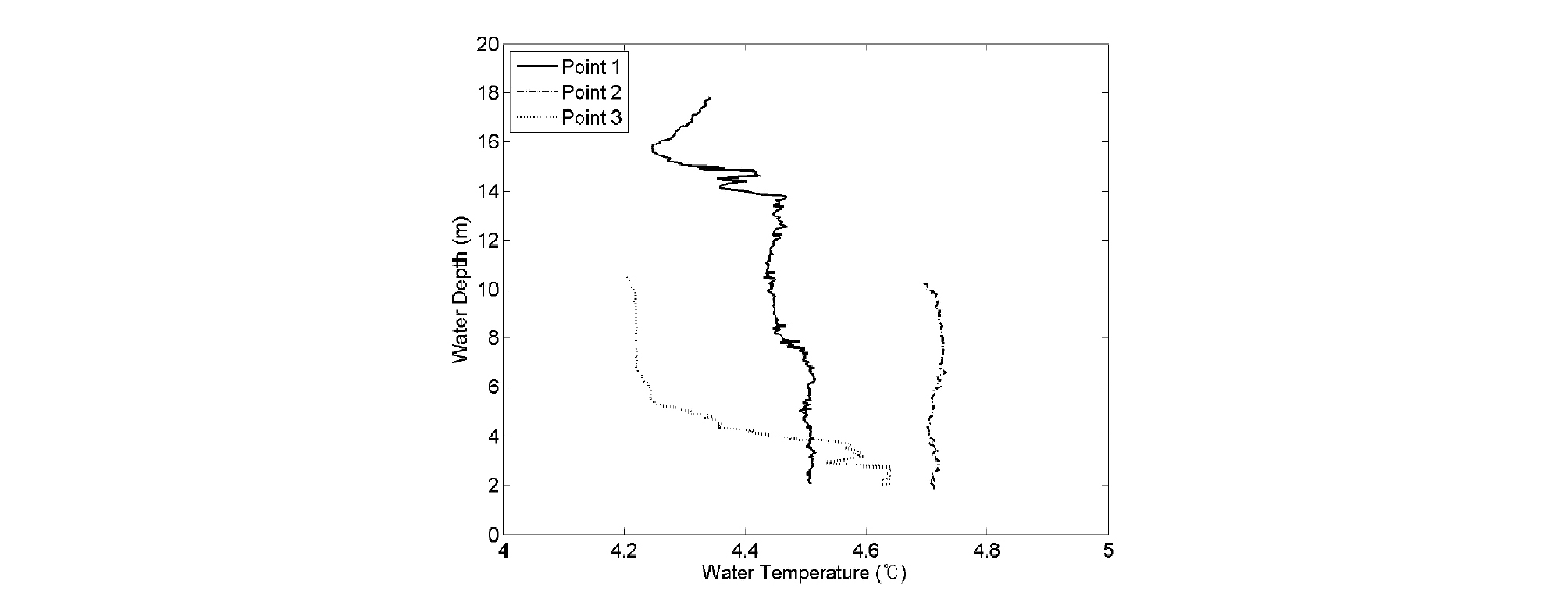
집중조사를 위해 선정된 평정저수지는 계곡지의 형태를 가지는 전형적 농업용 저수지이다. Fig. 2(e)에 나타난 지점 3은 수심이 18 m인 지점으로 저수지의 가장 최심부에 해당한다. 2015년 12월 9일, 27일, 2016년 1월 3일, 2월 23일 등 총 4회에 걸친 동계 조사 모두에서 최심부인 지점 3의 바닥면에서 어군이 형성되는 것을 확인하였다. 해당 일자에 수표면 온도는 저수지 전체를 통해 거의 일정한 값을 보였으며 지점 3을 제외한 저수지 전역에서는 어군의 형성이 잘 발견되지 않았다. Fig. 5는 2016년 2월 23일에 조사된 지점별 수온의 연직 분포를 나타낸다. 그래프에서 나타나는 것과 같이 동계의 수온은 바닥면으로 갈수록 오히려 증가하는 것을 알 수 있다. 즉 1℃ 이내의 미세한 온도 차이라고 하더라도 극한기에는 어류가 가장 수온이 높은 위치를 찾아 먹이활동을 줄이며 월동하는 사실을 알 수 있다. 동면을 하지 않는 외래어종의 특성을 고려할 때 수온을 따라 움직이며 서식처를 형성한다는 점도 확인되었다. 2016년 4월 6일에 시행된 현장조사에서는 저수지의 평균 수온이 약 13℃로 확인되었다. 저수지의 호안부에서 1년에서 2년생 사이의 작은 큰입배스의 먹이활동이 다수 확인되었으며, 수온이 약 13℃인 지점에서 채집된 외래어종을 살펴본 결과 산란공이 열리지 않은 것 또한 육안으로 관찰하였다. 2016년 6월 8일에 시행된 현장조사에서는 저수지 연안부의 수심이 얕은 곳 자갈 바닥지점에서 갓 부화한 큰입배스의 치어를 육안으로 관찰할 수 있었다. 큰입배스는 수컷이 산란장을 마련하며 산란 후 수정된 알과 부화된 치어를 숫놈이 적극적으로 보호하는 특성을 갖는 것으로 알려져 있는데, 갓 부화한 큰입배스의 치어 부근에서 알자리를 지키는 수컷큰입배스 또한 육안으로 관찰하였다. 큰입배스가 수온이 약 15℃에서 17℃ 정도가 되는 4월부터 5월까지의 기간 동안 산란하는 것을 생각해보면, 6월 초에 관찰된 큰입배스의 치어는 시기상 2차 산란인 것으로 추정해 볼 수 있다. 4계절에 걸친 현장조사를 통해 큰입배스가 형성한 서식처가 수온과 매우 밀접한 관련을 가지는 것을 확인하였다.
3.2 실험실 실험
앞서 수행된 현장실험 조사 결과로부터 외래어종의 서식처 형성과 수온이 상관관계가 있음을 확인하였으며, 이를 바탕으로 계절별 수온에 따른 외래어종의 서식처 특성을 보다 통제된 실험실실험 조건에서 살펴보았다. 실험 진행과정에서 수온을 제외한 다른 조건은 모두 일정하게 유지하였다. 수조에 설치된 히터와 냉각기를 이용하여 수온을 조절하였으며, 고수온대(수조의 좌측 1/3 구간), 중수온대(수로의 중앙 1/3 구간), 냉수온대(수로의 우측 1/3 구간)의 영역별 수온차가 3.0℃ 이내가 되도록 하였으며, 이식된 외래어종과 토착어종이 수조 내 공간에서 수온만의 영향을 받으며 서식처를 형성하도록 실험조건을 구축하였다. Table 1은 동계, 춘계, 하계에 수행된 실험실 실험에서 실험 조건으로 적용한 구간별 평균 수온을 나타낸다.
Table 1. The average water temperature in three different zones from winter to summer
| Area | Water temperature (℃) | ||
| Winter | Spring | Summer | |
| High temperature area | 14.1 | 17.4 | 27.2 |
| Moderate temperature area | 13.0 | 16.4 | 25.9 |
| Low temperature area | 11.6 | 14.8 | 23.9 |
동계에는 외래어종 및 토착어종 모두 비교적 온도가 높은 온수대에 서식처를 형성하는 것을 확인하였다. 이는 겨울철에 큰입배스가 보다 수온이 높은 곳을 찾아 이동하는 것을 확인한 현장조사 결과 및 이전의 연구결과와 일치한다(Karchesky and Bennett, 2004). 이 때 중수대와 온수대의 수온은 각각 14.1℃ 및 13.0℃로, 약 1.0℃의 수온차가 동계의 서식처 형성에 영향을 미침을 알 수 있다.
동계에서 춘계로의 계절 변화에 따라 가장 수온이 낮은 냉수대의 온도가 동계 실험 시 서식처를 형성하였던 온수대 온도와 동일해지도록 조절하였으나, 이식한 어류 모두 기존에 형성된 서식처에서 머무르는 현상을 확인하였다. 동시에 큰입배스의 산란을 관찰하였는데(Fig. 6(a)), 큰입배스의 산란은 형성된 서식처에 적응하였음을 나타내는 것으로 생각해 볼 수 있다. 일반적으로 큰입배스의 산란은 15℃ 이상의 수온에서 일어나는 것으로 알려져 있는데(Kohler et al., 1994), 산란이 일어난 때 온수대 영역의 수온은 약 17.35℃로 관찰되었다. 산란한 장소에서 알자리를 지키는 수컷 큰입배스를 꾸준히 목격할 수 있었다(Fig. 6(b)). 또한 산란기의 큰입배스는 외부의 자극에 예민하게 반응하는 것을 알 수 있었으며, 다른 개체를 위협하는 모습 또한 목격되었다. 알자리를 지키는 수컷 큰입배스 주변으로 나머지 큰입배스 또는 붕어의 서식처 형성은 불가능하였다. 온수대와 중수대의 수온 차를 약 1.0℃ 미만으로 조절할 경우에는 큰입배스가 중수대까지 서식처의 범위를 넓히는 반면, 구간별 온도차이가 1.0℃ 이상일 경우 뚜렷하게 온수대에 서식처를 형성하였다.

춘계에서 하계로의 계절변화에 따라 수조 내 수온이 모든 영역에서 21.0℃ 이상인 때에는 기존에 서식처를 형성한 온수대 이외에도 중수대와 냉수대에서 머무르는 것을 확인하였다. 하계에 큰입배스는 주로 냉수대에 서식처를 형성하였다. 냉수대와 중수대 온도 차이가 약 1.0℃가 되도록 조정한 경우 춘계와 마찬가지로 중수대까지 서식처를 형성하였으며, 비교적 동계보다는 수온에 둔감하게 반응하는 특성을 보였다. 달라지는 수온에 따른 큰입배스의 서식처 형성 특징을 살펴본 결과에 따르면, 하계에 큰입배스는 약 20.0℃부터 24.0℃까지의 범위에 해당하는 수온 영역을 가장 선호하는 것을 알 수 있다.
수온과 서식처 형성의 상관관계 외 추가적으로 토종어종과 외래어종을 같은 공간에서 사육할 경우 서식처 분할여부와 먹이경쟁이 나타나는지 살펴보았다. 외래어종은 수조에 함께 서식한 붕어와는 따로 구분되는 서식처를 형성하지 않았으며 함께 생활하는 것이 확인되었다. Ko et al. (2008)의 연구에 따르면, 채집된 큰입배스의 소화관 내용물에서는 피라미 다음으로 붕어의 비율이 매우 높은 것이 보고된 바 있다. 따라서 본 실험연구에서 붕어와의 서식처 분급화 현상 또는 큰입배스의 붕어 포식이 발생하지 않은 이유는 비교적 붕어의 크기가 커 큰입배스가 섭취할 수 있는 크기가 아니었기 때문으로 생각해 볼 수 있다. 미꾸라지와 금붕어, 빙어와 같이 크기가 작은 개체는 모두 큰입배스에게 매우 쉽게 포식된 것 또한 확인하였다. 큰입배스의 먹이활동을 살펴본 결과에 따르면, 동일한 양의 먹이를 동계, 춘계, 하계에 공급하는 경우 동계에 가장 오랫동안 먹이의 양이 유지되는 것을 확인할 수 있었다. 이는 동계에는 먹이활동을 활발하게 하지 않는 것으로 알려진 큰입배스의 서식특성과 일치한다. 먹이가 부족한 경우 비교적 큰 개체가 작은 개체를 잡아먹는 동종간 공식 현상이 발견되었으며, 블루길을 포식한 것 또한 확인하였다. 섭식 가능한 먹이가 부족한 환경에서 큰입배스는 먹이경쟁으로 인해 동종 간 공식이 발생하는 것으로 알려져 있으며(Hossain et al., 2013), 우리나라의 하천과 저수지에서 적응한 큰입배스의 공식 현상 또한 확인된 바 있다(Park et al., 2019).
큰입배스의 개체 크기에 따른 서식처 형성 특성을 살펴본 결과에 따르면, 큰입배스는 동종일지라도 개체의 크기에 따라 서식처를 달리 형성하는 것이 확인되었다. 또한 동종간 공식 현상이 나타나는데, 작은 개체들은 목숨에 위협이 되는 큰 개체를 피해 서식처를 형성하여 크기별로 형성되는 서식처의 위치가 다름을 확인할 수 있었다. 작은 개체들이 서식처를 형성하는 곳에는 은신처가 될 수 있는 좁은 돌 틈 또는 장애물이 있는 지역으로 나타났다. 보다 명확한 확인을 위해 작은 개체에게 위협이 되는 것으로 여겨지는 큰 개체를 제거하자, 작은 개체가 기존의 큰 개체가 머물던 자리에 서식처를 형성하는 것이 확인되었다.
형성된 서식처 내 장애물 부근에서 비교적 작은 크기의 큰입배스가 머무르는 것을 바탕으로 수온 이외에 장애물의 선호에 관한 실험을 추가적으로 수행하였다. 장애물 반응에 관한 실험에서는 환경 조건이 모두 동일한 수조에서 서로 다른 세 종류의 장애물을 설치하였다. 수조의 전면부 방향으로 장애물을 설치하여 큰입배스가 수조 밖 자극에 노출되지 않도록 설치한 경우와 큰입배스의 전면 시야를 차단하기 위한 방향으로 장애물을 설치한 경우, 상부 자극을 차단하는 장애물을 설치한 경우를 살펴보았다. 실험 결과로부터 큰입배스가 상부 자극을 차단하는 장애물을 가장 선호하였으며, 서식처 또한 상부 자극을 차단하는 장애물 아래 형성함을 확인하였다. 장애물과 수온의 두 가지 독립변인 중 큰입배스의 서식처 형성에 있어서 더욱 선호되는 인자를 찾기 위한 실험실 실험에서는 장애물이 더욱 선호되는 것을 확인할 수 있었다. 이러한 실험 결과는 Bevelhimer (1996)의 연구결과와 일치한다. Bevelhimer (1996)는 작은입배스(Smallmouth Bass)의 서식처 형성에 물리적 구조물, 먹이, 수온이 미치는 영향의 중요도를 살펴보기 위한 실험실 실험을 수행하였는데, 실험 결과로부터 작은입배스의 서식처 형성에 있어 가장 중요한 요인이 장애물의 유무인 것을 확인하였다(Bevelhimer, 1996). 이외에도 실험실 실험 및 현장조사로부터 큰입배스가 서식처 형성 과정에서 장애물 주변부를 선호하는 특성을 나타내는 것이 다수의 연구로부터 확인된다(Hubert and Lackey, 1980; Haines and Butler, 1969). 큰입배스가 장애물 부근에 서식처를 형성하는 원인으로는 빛과 포식자로부터 자신을 보호하기 위한 목적, 비교적 풍부한 먹이(Bevelhimer, 1996) 등이 제시되며, Savino and Stein (1989)은 큰입배스가 먹이의 종류를 구분하는 것에 근거하여(Nyberg, 1971) 장애물 아래에서 사냥할 먹이를 기다리는 것이라 주장하였다.
태양광선을 대신하기 위해 수조의 윗부분에 설치한 조명을 이용하여 조명 유무에 따른 서식처 형성 특성을 확인하기 위한 실험실 실험을 수행하였는데, 큰입배스는 수조 전역에 걸쳐 상단에 설치된 조명의 아래에는 머물지 않는 것으로 확인되었다. 이미 서식처가 형성된 영역에 조명을 비출 경우에는 조명을 피해 서식처를 옮기는 것 또한 확인되었다. Johnson (1993)의 연구에서는 장애물과 빛의 세기에 따른 서식처 형성을 살펴보았는데, 장애물의 선호도에 따른 서식처가 형성된 이후에도 광도에 따라 서식처를 옮기는 모습을 관찰하였다. 이를 바탕으로 외래어종의 서식특성에 장애물의 유무나 형태보다는 광도가 더 지배적인 영향을 미친다고 주장하였다(Johnson, 1993).
4. 결 론
본 연구는 우리나라의 민물생태계에 존재하는 큰입배스의 서식처 형성 특성을 살펴보는 것을 목적으로 수행되었다. 현장조사로부터 큰입배스의 서식처 형성과 수온이 밀접한 관련이 있음을 확인하였으며, 이를 바탕으로 수온에 따른 서식처 형성특성을 살펴보는 실험실 실험을 수행하였다. 4계절에 걸쳐 수행된 실험실 실험 결과로부터 큰입배스가 동계에는 비교적 수온이 높은 곳에서 서식처를 형성하며, 하계에는 수온이 낮은 곳에서 서식처를 형성하는 것을 확인하였다. 동계에서 춘계로 계절이 변화하는 때에는 온도의 변화와 무관하게 기존에 형성된 서식처에서 산란이 일어나는 것을 확인할 수 있었다. 수컷이 알자리를 지키는 큰입배스의 서식특성 또한 실험실 실험 수행 과정에서 관찰할 수 있었다.
수온에 따른 서식처 형성 외 토종어종과 외래어종의 종에 따른 서식처 분할 유무를 살펴보았는데, 붕어와 큰입배스가 동일한 서식처에서 함께 생활하는 것으로 나타났다. 이를 통해 개체의 크기가 유사한 경우에는 종에 따른 서식처 구분이 일어나지 않음을 확인하였다. 토종어종과 달리 큰입배스의 비교적 작은 개체와 큰 개체는 구분된 서식처를 형성하는 특성을 갖는 것으로 나타났다. 작은 개체가 큰 개체로부터 구분된 곳에서 장애물 부근에 머무는 것을 바탕으로 장애물에 따른 서식특성을 살펴본 결과, 전면 시야보다 상부 자극을 차단하는 장애물을 선호하는 것으로 확인되었다. 장애물과 수온, 조명의 유무가 서식처 형성에 미치는 영향을 살펴본 결과에 따르면 장애물에 대한 선호도가 가장 높았으며, 이미 서식처를 형성하였더라도 빛의 유무에 따라 서식처를 변경하는 것이 확인되었다. 그러나 장애물과 조명의 유무에 대한 실험실 실험은 비교적 간단한 장비 조작을 통해 이루어진 것으로 보다 향후 면밀한 실험실 실험이 수행될 필요가 있다고 판단된다.
4계절에 걸친 큰입배스의 먹이활동을 살펴본 결과에 따르면, 동계에는 먹이활동을 활발하게 하지 않으며 먹이가 부족할 때에는 동종간 공식이 발생하는 것을 확인하였다. 향후 수자원확보를 위해 수공시설물이 건설된다면 그로 인한 큰입배스의 확산은 더욱 활발해질 것으로 생각해 볼 수 있다. 이에 따라 장기적으로 큰입배스의 서식처 확산을 제어하는 방법이 요구되며(Hong and Son, 2003), 본 연구의 실험 결과가 기초자료로 활용될 수 있을 것으로 생각된다. 그러나 실제 하천의 환경은 매우 다양한 변수를 포함하고 있어 실험실 실험이나 분석의 과정에서 파악하지 못한 다양한 양상이 나타난다. 예를 들어, 이전의 연구 사례에서는 외래종의 감소로 인해 또 다른 형태의 생태계교란이 발생하는 것이 보고된 바 있다(Maezono et al., 2005). 따라서 실제 현장에서의 적용이 가능한 구조물의 개발과 이를 이용한 실험이 필요할 것으로 생각된다.
