

1. 서 론
2. 관련 연구
3. 조사구간에서 식생의 이입 현상
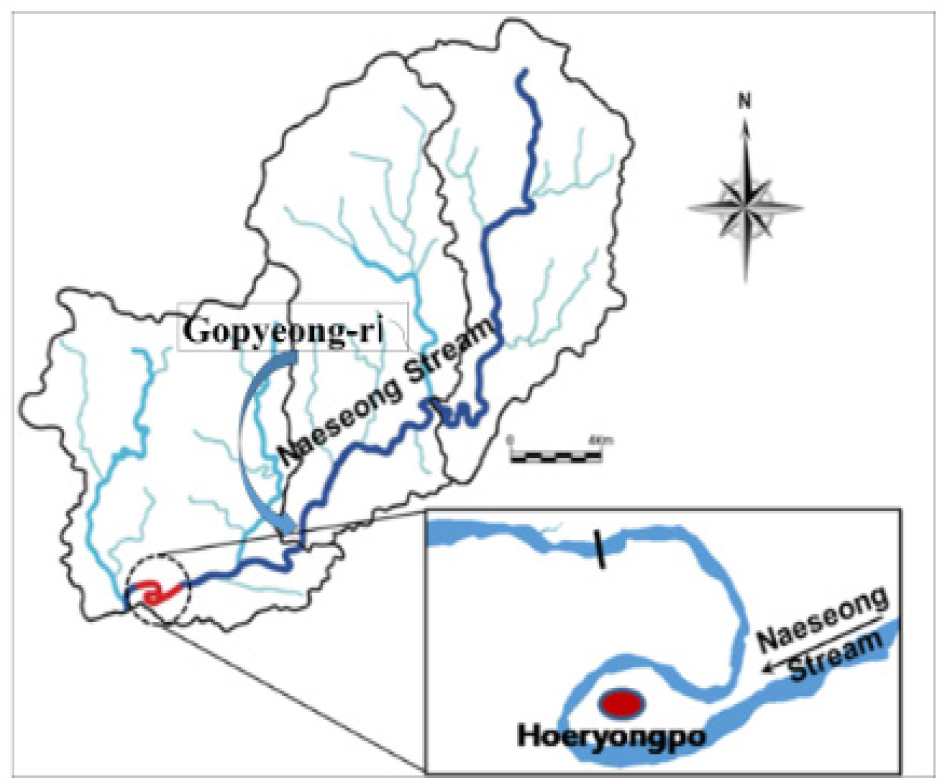
3.1 유역 및 하천 특성
3.2 식생 이입
3.3 식생 특성
4. 흐름 및 하상소류력의 수치해석
4.1 수치모의
4.2 모형의 검증
4.3 홍수사상 및 계산
5. 연구 결과 및 토의
5.1 식생생존의 한계조건
5.2 무차원 소류력 모의 결과
5.3 결과의 토의
6. 결 론
1. 서 론
국내의 많은 하천이 인간의 간섭이 있든 없든 이른바 ‘화이트리버’에서 ‘그린리버’로 바뀌고 있다(Choi et al., 2005; Egger et al., 2012; Woo et al., 2014). 이러한 현상은 국내에 국한된 것이 아니라 일본, 유럽, 미국 등 많은 지역에서 공통적으로 나타나고 있다(Asaeda et al., 2013; Garófano-Gómez et al., 2013; de Jalón et al., 2020). 이러한 하천내 식생이입은 생태적 영향(Gurnell, 2014)뿐만 아니라 강턱 침식, 홍수위 상승 등 공학적 문제를 일으킬 수 있다(Kiss and Fehérváry, 2023). 지금부터40년 전 흐름과 식생간 상호작용에 관한 선구자적 연구(Williams and Wolman, 1984) 이후, 많은 연구자들이 야외와 실험실에서 흐름과 유사이송, 지형 및 식생간 상호작용에 관한 조사연구를 수행하였고, 나아가 해석적 방법과 수치해석 등으로 관련 인자들 간 다양한 상호작용을 개별적, 집합적으로 모의하였다. 이러한 연구성과들은 2010년대 중반 이후 ‘state-of-the-art’ 총설 연구 논문 등에 잘 나와 있다(Gurnell et al., 2016; Solari et al., 2016). 그들은 공통적으로 하천에서 식생 이입에 기본적인 필요조건으로 흐름과 토양수분을 들고 있다.
본 연구는 하천에 식물의 씨가 사주에 정착한 후 발아하여 생장하는 이입 단계에서 기본적으로 요구되는 물리적 안정성에 초점을 맞춘 것이다. 즉, 실제 하천에서 관찰한 결과와 하천 흐름해석을 통해 얻어진 값들을 비교하여 식생의 유식물기(seedling) 및 유년기(juvenile) 등 이입기(recruitment stage)에 물리적으로 세굴(scour), 뿌리 뽑힘(uprooting), 매몰(burial) 등으로 사멸하는 흐름의 한계조건을 추정하는 것이다. 나아가 이 분야 연구에서 실험적, 관찰적 확인 없이 식생의 사멸조건으로 이용되는 현 기준을 재평가하는 것이다. 이를 위해 국내 하천에서 흔한 초본류인 명아자여뀌(node smart weed, 학명 Persicaria nodosa; Fig.1)를 대상으로 연구하였다. 갈대나 달뿌리풀이 국내에서 가장 한 초본류지만 이는 다년생이고 씨보다는 뿌리를 통해 번식하므로 상대적으로 관찰, 분석 하기 어려워 단년생인 명아자여뀌를 선택하였다.

본 연구에서 선정한 대상하천구간은 회룡포라 불리우는 낙동강 제1지류 내성천 구간이다. 내성천은 국내 전형적인 모래하천 중 하나이며, 이 중 회룡포 구간은 내성천 최하류에 위치한 하도가 거의 180도로 구부러지는 곳이다(Fig. 2). 이 구간은 이 지역의 오래된 항공사진(1965년) 관찰 결과 2013년까지 흰 모래밭이 거의 그대로 유지된 이른바 ‘화이트리버’ 이다(Fig. 3).

본 연구에서는 2013, 2014, 2015년 세 연도에 걸쳐 매년 조사구간 하안에서 명아자여뀌의 생장을 관찰하였다. 구체적으로, 4월에 발아하여 가을에 수분 후 시들 때까지 식물의 무게와 키 높이를 측정하였다. 동시에 약 한 달 간 씨앗 퍼짐도 관찰하였다. 다음 여러 연구를 통해 검증된 하천수치모형을 이용하여 해당구간에서 흐름과 하상소류력을 계산하여 매년 측정된 초본류의 생장 및 사멸 기록과 비교하였다. 본 연구의 조사대상 초본류인 명아자여뀌는 그 전 해 가을에 생성된 씨앗이 수매작용(hydrochory)을 통해 하안에 정착한 후 그 해 4월에 발아한다.
2. 관련 연구
내성천은 2010년대부터 식생과 흐름-유사이송 관련 몇 연구가 진행되었다. Lee et al. (2019a)은 내성천을 대상으로 2013-2018년 간 6년 동안 비교적 장기적으로 수문, 식생, 지형 등 다학제적 조사연구를 수행하였다. 연구 결과 내성천의 식생이입/활착은 특히 고평리 구간(Fig. 2)에서 1987년부터 서서히 시작되었으며(Fig. 4), 2013년 이전에는 하류 구간에서, 그 이후 2015년부터 전체 구간에서 식생이 활착하였다. 그들은 특히 2015년 강수량 자료에 주목하여 이는 1982년 다음으로 가장 적은 값으로서, 여름철 첨두유량은 50년 이래 최저치를 보였다. 이로 인해 하천에 식생 이입/활착에 유리한 물리적 조건이 조성되었음을 강조하였다. 그들은 이은 논문에서(Lee et al., 2019b) 내성천 식생은 일부 지점에서 홍수로 인해 사멸되어 사주는 다시 ‘나지’가 되었으나 여러 구간은 목본으로 천이 되었음을 강조하였다. 결론적으로 그들은 1990년대 이후 내성천에 나타난 식생이입 및 하도변화, 그에 따른 하천경관 변화는 주로 특정 시기의 하천유량 감소로 인한 것으로 추론하였다.

Fig. 4.
Yearly-changes in surface cover of the Naeseong-cheon Stream near Gopyeongri (Source: Fig. 1, Lee et al., 2019b)(Green color is vegetation)
나아가 Lee et al. (2019c)은 내성천과 서천 합류점의 직하류에 있는 만곡부를 대상으로 1980년부터 2016년까지 비조절 상태에서 사주상 식생이입 상황을 조사하고 2차원 수치모형을 적용하여 분석하였다. 그 결과 내성천과 같은 비조절하천에서도 1987년 이후 식생이입이 시작되어 2016년 기준 식생역은 수역을 제외한 전체 하천구간 면적의 70% 정도를 차지하였다. 그들은 이 연구에서 6-7월 홍수 기간 중 첨두홍수량과 해당 기간의 식생역 면적은 R2 = 0.73의 상당한 역상관 관계가 있음을 보이면서 식생이 사주에 이입되어 발아하여 유식물기, 유년기로 성장할 때 강한 홍수가 있으면 하상소류력에 의해 초본류는 사멸된다는 가설을 제시하였다. 마지막으로 Woo et al. (2020)은 내성천 회룡포 하천구간을 대상으로 2차원 수치모형을 적용하여 식생 이입 및 사멸/생존 조사기간을 포함한 전후 연최대 홍수사상에 대해 흐름 및 하상소류력 분포를 분석하여 해당 초본류 식생에 대한 사멸 및 생존 조건을 간접적으로 제시하였다.
본 연구는 이와 같은 Woo et al. (2020) 연구방법론을 적용하여 같은 구간에 대해 1) 조사구간에 적용할 수치모형을 해당 하천 자료를 이용하여 검증하여 계산결과의 신뢰도를 높이고, 2) 조사기간의 홍수사상을 2013-2015년 식생조사기간으로 국한하여 초본류 사멸/생존 조건의 도출범위를 좁히고, 3) 이 결과를 기존의 다양한 사멸/생존 조건 가정치와 비교 평가하는 것이다.
3. 조사구간에서 식생의 이입 현상
3.1 유역 및 하천 특성
내성천은 낙동강의 제1지류로서, 하천길이 108.2 km, 유역면적 1,806.7 km2, 하상재료의 평균입경은 1.6 mm, 하도경사는 9/10,000~1.7/1,000이다. 하곡은 비교적 좁아서 그 넓이는 하폭의1.5~3.0 정도이며, 하도의 측방변동은 하곡의 고지와 구릉으로 제한되어 있다. 유역의 연평균 강수량은 1,231 mm 이며, 연도별로 큰 변화는 없다. 내성천은 상류에 영주댐이 건설되어 담수를 시작하기 전인 2019년까지 사실상 비조절하천, 즉 비교적 자연상태의 유황을 보인 하천이었다. 따라서 본 연구조사 기간인 2013-2015년은 비조절상태이었다. 조사 구간의 2년빈도 홍수량은 일유량 기준 690 m3/s 이다.
3.2 식생 이입
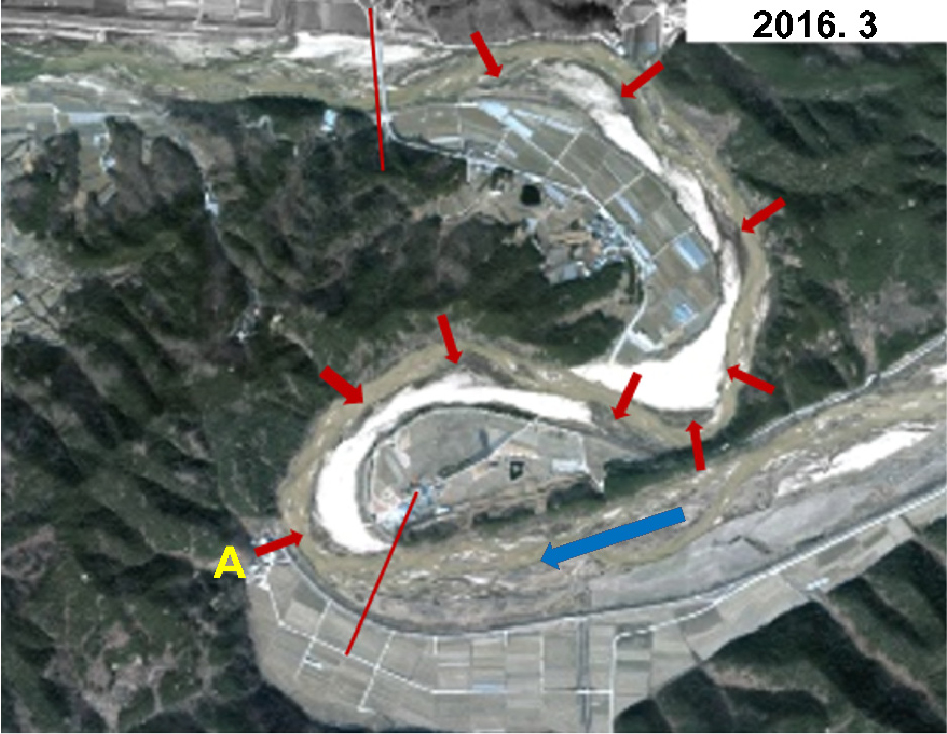
Fig. 3은1965년부터 2017년까지 조사구간의 식생이입 상태를 보여주는 항공사진이다. 식생은 대부분 초본류인 갈대와 달뿌리풀, 그리고 명아자여뀌이며, 일부 버드나무가 패치형태로 있다. 이 그림에서 보는 바와 같이 조사구간에서 ‘화이트’ 사주는 적어도 2013년 12월까지는 그 상태를 유지한 것으로 보인다. 그러나 2016년부터 사진에서 보이는 첫번째 만곡부의 하류 내측부에 식생이 나타나기 시작하여 그 하류 하안에 패치형태로 연속적으로 나타났다. 현지 조사 결과 저수로 인근 하안식생은 대부분 명아자여뀌 이며, 다만 사주상 비교적 높은 것의 식생은 달뿌리풀이다. 명아자여뀌는 ‘맨’ 사주에 이른바 개척종 식물로 자주 나타나는 것으로 알려져 있다. Fig. 5는2016년 3월 사진을 확대한 것으로서 그 전 해 하안에 이입된 식생패치를 잘 보여준다.
3.3 식생 특성
조사 결과 본 연구의 대상 종인 명아자여뀌는 통상 늦은 4월에 발아를 시작하여 늦은 6월에 30 cm정도 자랐다. 이 때 줄기의 직경은 4 - 9 mm 정도 이었으며, 뿌리는 26 cm 정도 자랐다. 지상부의 건조중량은 3.9 g, 뿌리부는 0.8 g정도 이었다. 본 연구의 관심 생장단계도 이 같은 씨앗의 유년기 이입단계이다. 명아자여뀌는 늦은 8월 최대 성숙기를 지나서 식물 높이는 평균 50 cm 정도, 뿌리 길이는 45 cm, 줄기 직경은 13.2 mm, 지상부의 건조중량은 75.6 g, 지하부는 11.5 g이었다. 이 정도의 뿌리 생장 상태에서는 어느 정도 큰 홍수가 오더라도 사주 세굴이나 매몰 등에 의해 쉽게 사멸되지 않을 것으로 추정된다. 그 후부터 수분을 시작하여 씨를 퍼뜨리고 갈색으로 시들기 시작하였다.
Fig. 6은 2013, 2014, 2015년 3년간 각 시기별 Fig. 5의 방형구 A지점에서 명아자여뀌 생장을 보여준다. 이 그림에서 보는 바와 같이 2013년에는 4월 상순에 발아하여 5월 상순에 7 cm 높이까지 자라다가 7월 이전에 사멸하였다. 반면에 2014년과 2015년 식생은 각각 평균 50 cm까지 자라고 9월 중순부터 키가 점차 줄어들다가 늦은 가을에 시들기 시작하였다. 여기서 연구자들의 관심은 2013년 6월 홍수로 식생이 사멸한 반면에 2014년 홍수에 식생이 생존한 점이다. 2015년에는 특별한 홍수가 없었다.
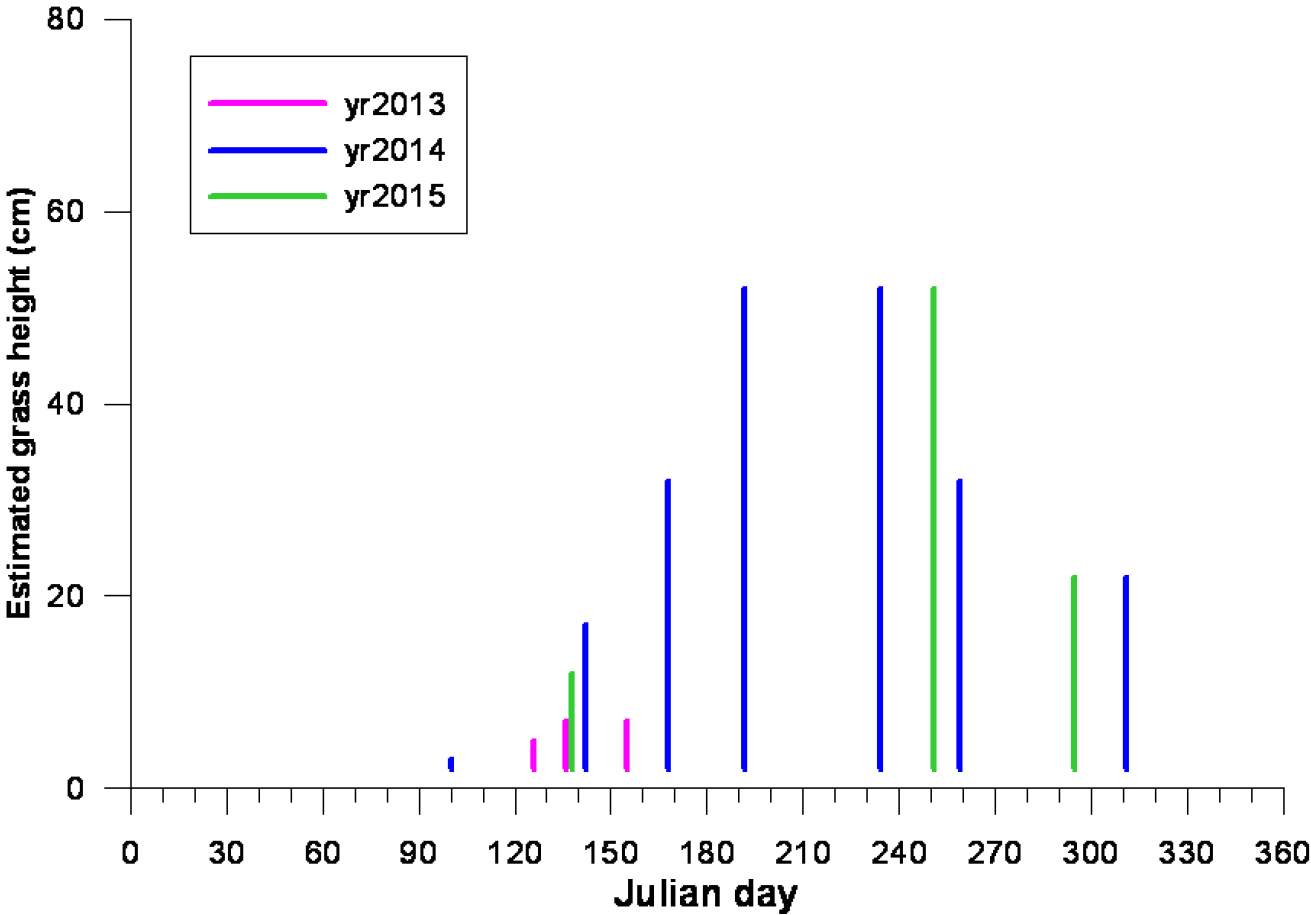
Fig. 6.
Changes in plant heights at study transect “A” in Fig. 5 with time for the years of 2013, 2014, and 2015
Fig. 7은 같은 지점에서 각각 2013, 2014, 2015년에 찍은 사진들이다. Fig. 7(a)에서 보는 바와 같이 Fig. 5의 A 지점에서 2013년 8월에 하안사주는 식생이 전혀 없는 ‘화이트리버’ 였다. 그러나 2014년 9월에 찍은 사진에는 하안사주에 이미 명아자여뀌가 이입된 것으로 나타났으며, 2015년 9월에는 더 조밀하게 활착된 것으로 나타났다. 이러한 변화는 그 그간의 홍수와 밀접한 관계가 있으며, 이에 대해서는 다음 장에서 구체적으로 논의한다.

Fig. 7.
Series of photos of the study transect “A” in Fig. 5(a) (Photo (a) Taken on August 15 2013, (b) Taken on September 4 2014, (c) Taken on September 24 2015). (Node smartweeds not seen in photo (a), while they were recruited along the riparian shore in photos (b) and (c))
4. 흐름 및 하상소류력의 수치해석
4.1 수치모의
본 연구에서 조사구간의 하천흐름 및 하상소류력 특성을 분석하기 위해 iRIC모형에 장착된 Nays2DH 모형(Shimizu et al., 2020)을 이용하였다.이 모형에 대해서는 같은 저자들이 최근에 게재한 연속논문(Woo et al., 2025a; 2025b)에 자세히 소개되어 있으므로 그 자료를 참고할 수 있을 것이다.
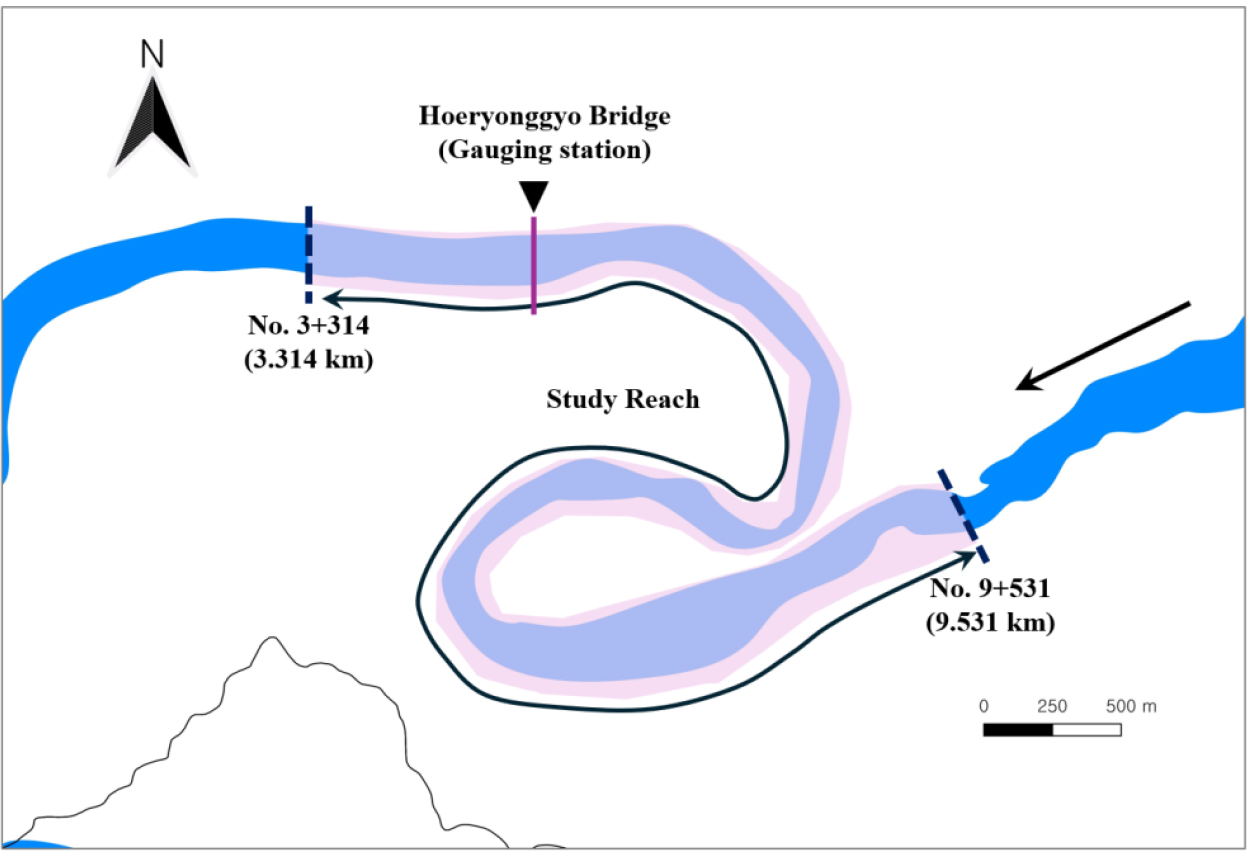
본 연구에서 조사구간은 Fig. 5의 적색으로 표시된 상류단 선과 하류단 선 사이 구간이다. 구간 거리는 3.8 km 이며 하폭은 209~406 m 까지 변한다. 하상경사는 평균 0.0009이며, 하상재료의 평균 입경은 1.5 mm로서 굵은 모래이다. 그러나 모형 적용구간은 조사구간보다 더 크게 책정하여, Fig. 8과 같이 회룡포 수위관측소(경상북도 예천군 용궁면 대은리 950, 36°35'10.0"N 128°19'10.0"E)를 포함하여 총 6.2 km 이다. 모형적용을 위해 하천기본계획(MLTM, 2013)에서 제시한 지형자료를 이용하여 계산구간내에 205×25(=5,125)의 계산격자망을 구축하였다. 계산의 정확도를 높이기 위해 만곡부와 협착부는 조밀하게, 그 밖에 구역은 상대적으로 느슨하게 망을 구축하였다. Manning계수는 Fig. 5의 항공사진을 참고하여 식생이 없는 저수로나 사주에는 하천기본계획(MLTM, 2013)에서 제시한 0.031을, 식생이 있는 홍수터에는 0.035를 설정하였다(Chow, 1959). 그러나 해당 식생은 모두 초본류이므로 목본류 식생에 의한 추가적인 저항은 고려하지 않았다. 계산시간 간격은 0.01초로 설정하였다.
4.2 모형의 검증
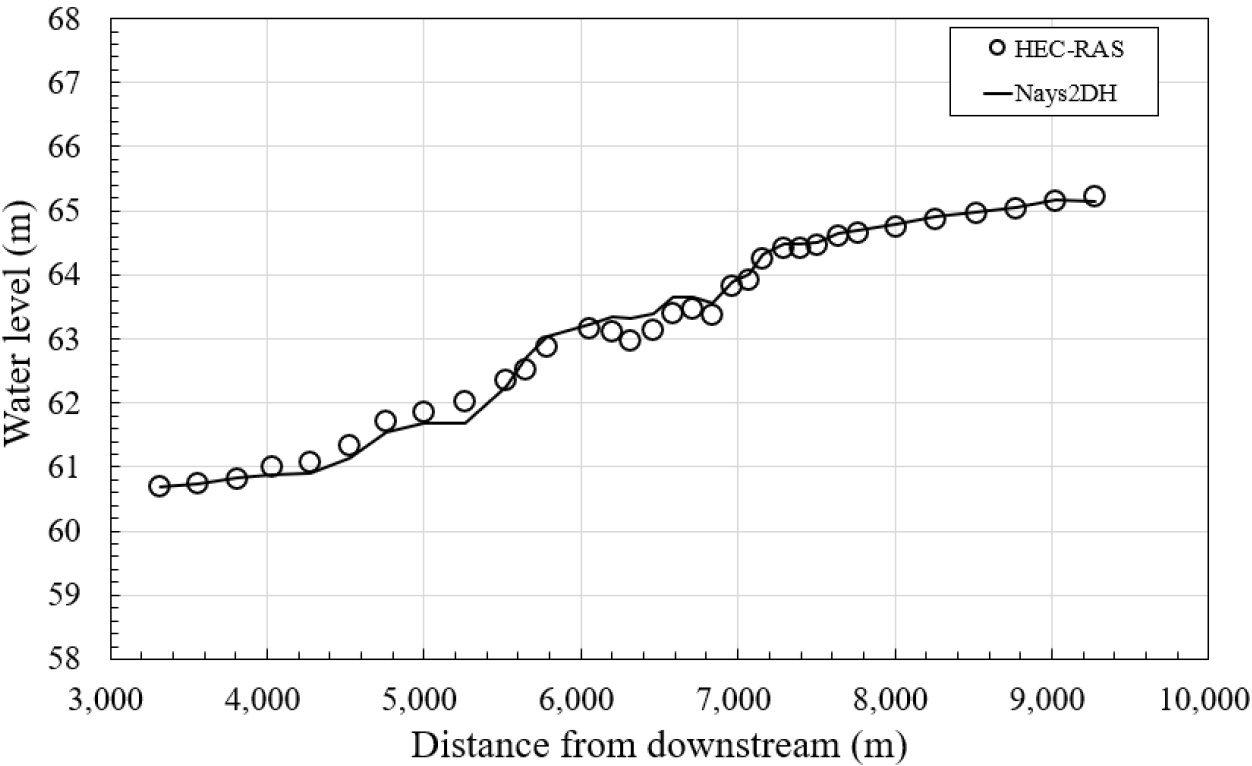
2차원 수치모형의 적용성을 검증하기 위하여, 계산구간에서 계획홍수량(100년빈도 유량)인 3,580 m3/s을 적용하여 부등류 수치모의를 수행하고, 그 결과를 1차원 수치모형인 HEC-RAS의 모의 결과와 비교하였다. 이 구간은 계획홍수량에 대한 실측 수위자료가 없기 때문에 국내외에서 널리 쓰이는 HEC-RAS를 기준으로 간접 검증하였다.
Fig. 9을 보면 1차원 모형의 결과와 2차원 모형의 결과가 비교적 잘 일치하고 있다. 그러나 급만곡부 구간인 5,000 m 구간에서 2차원 모형이 약간 적게 계산되고, 6,300 m 구간 에서 약간 크게 나타난다. 이는 2차원 모형은 만곡부에서 2차류의 흐름을 고려하여 모의하고 있으나 1차원 모형은 이러한 특성을 고려하지 않고 모의하기 때문으로 판단된다.
두 모형의 수치모의결과를 정량적으로 비교, 검증하기 위한 지표로 NSE (Nash-Sutcliffe Efficiency)와 RMSE (Root Mean Square Error)를 이용하였다. NSE는 Nash-Sutcliffe 효율로서 수치모의결과의 정확성을 검증하는 지표이며, 지표치가 1에 가까울수록 관측치와 수치모의 결과가 일치하는 것을 의미한다(Gupta and Kling, 2011). RMSE는 평균 제곱근오차로서 각 지점별 오차의 제곱을 산술 평균한 값들의 제곱근으로서 불일치도를 나타내는 지표이며, 지표값이 0에 가까울수록 일치하는 것을 의미한다(Chai and Draxler, 2014). NSE, RMSE의 산정식은 각각 Eqs. (1) and (2)와 같다.
여기서는 측정값, 는 측정값의 평균, 은 모의결과이다. 위 식을 이용하여 1차원 수치모의 결과와 2차원 수치모의 결과를 비교분석한 결과, NSEC는 0.989, RMSE는 0.15로 나타났으며, 두 값들이 서로 잘 맞는 것으로 나타났다.
4.3 홍수사상 및 계산
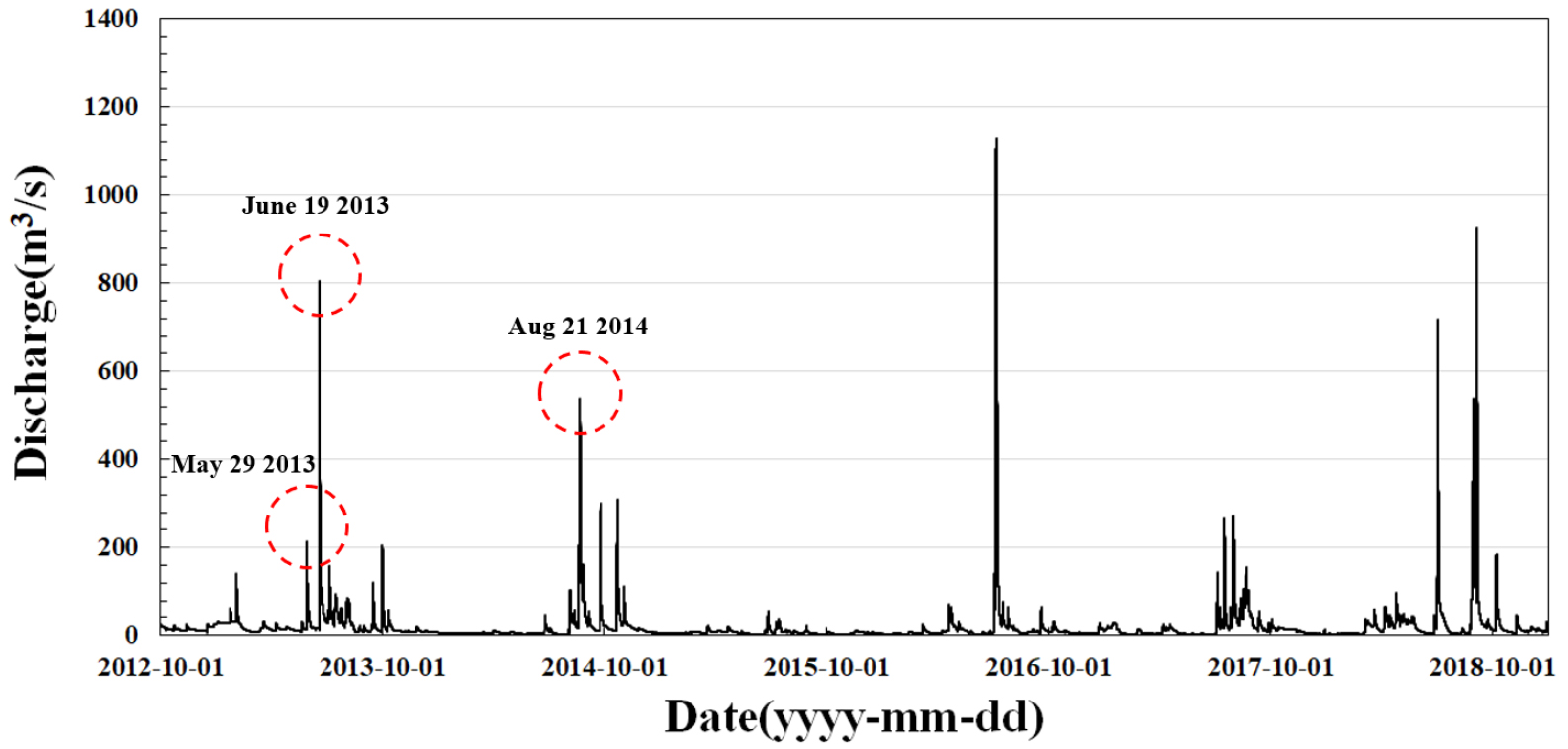
Fig. 10은 2012년부터 2018년까지 회룡포 수위관측소에서 일평균유량의 수문곡선을 보여준다. 이 구간에서 2년 빈도 홍수량은 690 m3/s이며, 이 그림에서 보는 바와 같이 7년 사이에 4번 정도 2년 빈도 홍수를 초과하는 홍수가 발생하였다. 본 연구에서는 2013, 2014, 2015년 식생이입이 주 관심 대상이므로 홍수사상도 그에 맞게 2013년부터 2015년까지 제한하여 Fig. 10에서 원으로 표시한 것과 같이 2013년에 2개 사상, 2014년에 1개 사상을 선택하였다. 2013년에 비교적 작은 홍수사상을 채택한 이유는 본 연구에서 2013년 봄-여름 계절이 임계상태로 보이므로 작은 홍수 사상에 대한 하상소류력을 검토하기 위함이다. 이를 정리하면 Table 1과 같다.
5. 연구 결과 및 토의
5.1 식생생존의 한계조건
전술한 바와 같이 하안사주 식생생존의 한계조건은 사주 표면의 물리적 안정성, 토양습윤(Gurnell et al., 2016), 식생의 침수 지속시간(Friedman and Auble, 1999; Calvani et al., 2019) 등으로 요약될 수 있다. 그러나 모래하천 하안에서는 지하수위가 지표수위와 사실상 동일하게 거동하므로(Woo et al., 2011) 토양습윤은 대체로 보장된다. 또한 국내 초본류의 침수 허용기간이 대부분 2주 이상인 반면(Shin et al., 2012), 실제 홍수에 의한 연속 침수기간은 보통 1주 미만이므로, 이 조건 또한 초본류 생존의 주요 한계요인이 되지 않는다. 따라서 본 연구에서 보다 중요한 요소는 하상의 물리적 안정성, 즉 세굴, 뿌리뽑힘, 매몰과 같은 물리적 불안정성에 대한 식생의 한계조건이다.
그러나 초본류의 경우, 이러한 물리적 불안정성에 따라 식생이 사멸하는 조건을 원인 기작 별로 체계적으로 구분한 연구는 거의 없다. 이는 초본류가 목본류에 비해 홍수 시 물리적 안정성이 현저히 낮고, 종·성장단계·사멸기작의 구분이 상대적으로 불명확하기 때문이다(Bendix and Hupp, 2000; Corenblit et al., 2007).
구체적으로, 기존 연구자들은 수변식생의 물리적 사멸조건을 설정하기 위해 하상소류력 또는 무차원 하상소류력(Shields 수)을 사용해 왔다. Benjankar et al. (2011), Egger et al. (2012)은 전자를 활용하였으며, Deltares (2019), Suzuki et al. (2012), Lee et al. (2019c) 등은 후자를 적용하였다. 하상소류력 접근법은 식물종 및 성장단계별 세굴·뿌리뽑힘·매몰에 대한 한계소류력을 필요로 하나, 기존 관련 모형들은 실험적·현장적 근거가 아닌 일종의 경험측(rule-of-thumb)에 의존하는 경향이 있다. 그나마 이러한 정보는 초본류보다 목본류에 편중되어 있다. 무차원 하상소류력 접근에서도 종·성장단계·사멸원인별 Shields 수가 필요하지만, 이를 도출한 실증적 연구는 찾아보기 어렵다.
따라서 본 연구에서는 다양한 초본류 사멸 요인을 구분하지 않고 ‘lumped’ 된 인자로서 무차원 하상소류력, 즉 Shields 수를 이용한다. 참고로 하상 불안정성은 하상재료가 움직이기 시작한 이후 발생하므로, 물리적 불안정성에 의한 초본류 사멸조건에서 유사량·유사농도 등은 후차적 요인에 해당하며, 입경분포는 통상 중앙입경으로 한다.
하상에 미치는 무차원소류력, , 을 수학적을 표시하면,
위 식에서 𝛾는 물의 단위중량, 는 유사입자의 단위중량, D는 유사입자의 직경, 는 하상전단응력이다. 하상전단응력, 또는 하상소류력 는 다음과 같이 얻어진다.
위 식에서 d는 수심, S는 흐름의 에너지 경사이다.
사립자가 움직이기 시작하는 이른바 운동의 시작 조건은 Shields 수를 기준으로 보통 0.047(Miller et al., 1977) 이지만 이는 사립자의 형상, 입도분포 등에 따라 달라지기 때문에 넓은 범위에서 0.03-0.06 정도이다. 그러나 하안, 또는 사주 상 식생의 물리적 불안정성에 관한 직접적인 연구는 문헌상 찾아보기 어렵다. 다만 기존 관련연구를 보면 사립자 운동의 시작조건에 준하여 해당 하안이나 사주를 구성하는 사립자의 Shields수에 해당하는 0.047값을 그대로 적용하거나(Friedman and Auble, 1999), 조금 더 큰 값으로 0.05-0.06정도 사용한다(Woo et al., 2014). 한편, Bertoldi et al. (2014)은 Li and Millar (2011) 자료를 준용하여 부드러운 식생, 보통의 식생, 강한 식생 등으로 나누어 각각 0.13, 0.17, 0.21 등을 사용하였다. 그러나 이러한 수치들의 정확한 근거와 식생조건의 분류에 대한 구체적인 설명은 해당 문헌에 나와 있지 않다.
5.2 무차원 소류력 모의 결과
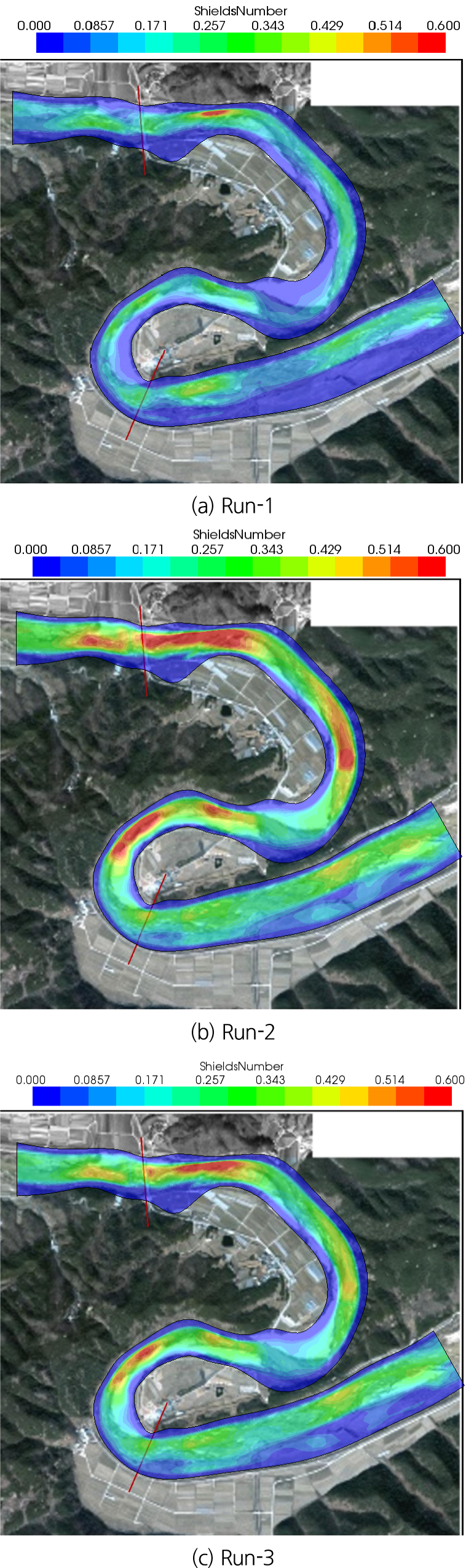
Fig. 11은 각 사상 별 해당년도의 항공사진에 계산된 Shields 수를 겹친 그림들이다. Fig. 11(a)에서 보는 바와 같이 대부분의 조사구간 사주에서 Shields 수는 0.085를 넘지 못한다. 다만 회룡교 상류에 하폭이 좁은 구간에서 Shields 수는 0.6 이상을 나타낸다. 그러나 Fig. 11(b)에서 보는 바와 같이 Run-2에서 회룡교 상하류의 하폭이 좁아서 흐름이 집중되어 유속이 빠르거나, 회룡포 급만곡부 외측에서 흐름이 집중되는 구간에서 Shields 수가 0.6 이상으로 나타났다. Run-3(Fig. 11(b))의 경우 Shields 수는 회룡교 상하류의 하폭이 좁거나 회룡포 급만곡부 외측의 흐름이 집중되는 곳에서 큰 값을 나타냈다.
5.3 결과의 토의
Fig. 5에 적색 화살표로 표시한 바와 같이 2016년 3월 항공사진에 보이는 각 식생 패치(지난 겨울에 죽은 식생) 별 Shields 수를 검토하기 위해 상류에서 하류방향으로 각각 A, a, b, …, h등으로 번호를 매겨 Fig. 12(a)에 나타냈다. 한편 비교를 위해 우측 Fig. 12(b)에 2013년 12월 항공사진을 표시하였다. 이 사진에서 구간 c는 하안사주 상 명아자여뀌 군락과 호안 가까이 타 식물군락이 혼재되어 있어 검토에서 본 제외하였다. 다음 각 사상별 해당 패치의 Shields 수를 Table 2에 나타냈다. 참고로 조사구간 내 모든 소구간(A, a-h 등 9개)에서 명아자여뀌는 일조량, 토양습윤 등 생장조건이 같으므로 생장상태도 사실상 같은 것으로 나타났다. 이에 따라 A 구간은 3년에 걸쳐 줄기/뿌리 생체량, 줄기 밀도, 높이, 사진촬영 등 생장상태를 정밀 조사하였고, 다른 구간은 정성적으로 관찰하였다.

Table 2.
Dimensionless bed shear stresses at sub-reaches marked with “A” and “a-h” in Fig. 12(a) for each run
Fig. 12(b)에서 보는 바와 같이 2013년 12월 당시 조사구간 하안에서 식생은 없었으며(즉 그 해 여름철 홍수로 사멸되었으며), 특히 Fig. 7(a)에서 보는 바와 같이 A 지점에서 그 해 8월에 없었다. 그러나 Fig. 6에서 보는 바와 같이 그 해 6월까지는 7 cm 높이의 식생이 있었다. 즉, 그 해 5월 29일 홍수에는 생존하였다는 것이다. 이 홍수로 인한 Shields 수는 A지점에서 0.11이고, 각 패치 별 0.11-0.19 범위이며, 평균 0.14이다. 그러나 그 해 6월 19일에 Run-2 홍수로 명아자여뀌는 모두 사멸하였으며, 이 홍수사상에서 Shields 수는 A 지점에서 0.24이며, 각 패치 별 0.24-0.52 범위 이며, 평균 0.36이다. 따라서 7 cm 높이의 2개월 된 유년기 명아자여뀌는 5-6월에 적어도 Shields수 0.19에서 생존하였으나, 0.24에서 물리적 불안정성으로 사멸한 것으로 추론할 수 있다. 0.19는 사립자의 운동시작 기준의 상한치인0.06보다 큰 값이며, Bertoldi, et al. (2014)나 Li and Millar (2011) 자료에 나오는 ‘보통의 식생’ 수준에 해당한다.
다음 본 식생 이입 및 생장 연구에서 중요 연도인 2014년을 보면 Fig. 6에서 보는 바와 같이 그 해에는 식생이 모두 지속적으로 성장하여 가을, 겨울에 자연적으로 시든 것으로 나타난다. 즉 식생은 2014년 8월 Run-3의 홍수에도 생존하여 성장하였다는 것을 시사한다. 또한 그 홍수 2주 후인 9월 4일에 촬영된 A 지점의 사진(Fig. 7(b))에 식생이 모두 관찰된다. 그 해 명아자여뀌는 봄에 발아하여 4개월 후 50 cm 성장하였다. 따라서 4개월 지나 성숙한 명아자여뀌는 Table 2에서 알 수 있듯이 적어도 7-8월에Shields 수 0.37에서 살아남은 것을 알 수 있다.
마지막으로, Fig. 10에서 보는 바와 같이 2015년에는 홍수가 없었으므로 그 해 명아자여뀌는 이입과 생장의 최적조건 이었음을 추론할 수 있다. 여기서 강조할 것은 본 연구에서 토양습윤은 하안식생을 대상으로 관찰하였기 때문에 명아자여뀌 생존의 한계조건이 되지 않는다는 점이다. 모래하천에서 하안이나 사주의 지하수위는 보통 하천 수위와 같다(Woo et al., 2011).
6. 결 론
본 연구의 조사구간에서 조사기간 전 후 약 50년 간의 항공사진 비교․분석, 3년에 걸친 현장조사, 그리고 조사기간 내 주요 홍수사상에 대한 2차원 수치해석 등을 통해 얻어진 결론은 다음과 같다.
모래하천에서 명아자여뀌와 같은 초본류는 식물의 유식물기 및 유년기 시절인 4-6월에 일정 규모의 홍수가 없는 경우 물리적 안정성이 보장되어 하안사주에 활착할 기회가 커진다.
7 cm 높이의 2개월 된 명아자여뀌는 무차원소류력 Shields 수 0.19에서 생존하나, 0.24에서 물리적 불안정성으로 사멸하는 것으로 나타났다. 따라서 명아자여뀌의 이입 초기단계(2개월 이전)에서 사멸조건은 Shields 수로 0.19-0.24 범위에 있으며, 이 값은 사립자의 운동시작 기준의 상한치인 0.06보다 훨씬 큰 값이다.
그러나 성숙기인 50 cm 높이의 4개월 된 명아자여뀌는 Shields수 0.37에서도 생존하는 것으로 추정된다.
사립자의 한계 Shields 수인 0.045, 또는 상한치인 0.06을 하상의 물리적 불안정성으로 인한 이입단계 식생의 사멸 기준으로 가정하는 현 하천식생 동역학 실무는 초본류 이입기의 경우 과소한 것으로 추정된다.
마지막으로, 본 연구는 명아자여뀌라는 국내에서 흔한 하안 초본류를 대상으로 식생의 이입기(유년기)에 초점을 맞추어 현장조사와 흐름해석 결과를 통해 식생 사멸조건을 무차원 하상소류력이라는 물리변수를 이용하여 추론한 것이다. 따라서 식물 종 별, 생장단계 별, 식생주변의 세굴, 뿌리 뽑힘, 매몰 등 다양한 사멸 원인을 구분하여 그 영향을 분석하기보다는 하상의 불안정성을 대표하는 간접적 지표로서 쉴즈 수를 이용하였다. 따라서 이러한 접근법의 내재적 한계를 넘기 위해서는 앞으로 식물종 별(초본, 목본 등), 식생의 생장단계(유식물기, 유년기, 성숙기) 별, 사멸조건의 기작(세굴, 뽑힘, 매몰 등) 별 세분화된 연구가 필요하다.
