

1. 서 론
식생은 하천환경을 구성하는 요소이며 수생태계의 건강성 유지를 위해 필수적 역할을 한다. 식생은 하천의 흐름을 변화시켜 유사의 퇴적, 침식에 영향을 미치며 저장대 형성을 통해 오염물질의 축적에도 영향을 미친다. 하도 내에 존재하는 식생은 수심과 식생 높이의 관계에 따라 정수식생, 침수식생, 부유성식생으로 구분할 수 있다(Yang, 2024). 정수식생은 항력을 일으켜 유속의 전단흐름을 감소시키는 것과 동시에 난류를 일으켜 오염물질의 전단혼합을 감소시키는 역할을 한다(Nepf et al., 1997). 침수식생 경계면에서는 흐름대와 식생대 사이에서 전단흐름이 강화되고, 대규모 와류를 일으켜 연직난류혼합을 증가시킨다. 이로 인해 부유사의 침강속도와 연직 혼합의 균형에 의해 식생과 흐름 경계면에 부유사가 정체하여 고농도의 부유사가 관찰되기도 한다(Li et al., 2020). 이와 같이 식생에 의해 변화한 난류 구조는 난류운동에너지(turbulent kinetic energy, TKE)와 난류의 특성길이(integral length scale) 변화에 영향을 미치며, 그 결과 난류에 의한 오염물질의 혼합은 식생이 없는 수로와 차이를 일으키게 된다. 식생에 의해 변화하는 난류운동에너지와 난류 특성길이의 곱은 난류확산계수와 비례하므로(Nepf, 1999), 난류확산계수의 변화로부터 식생에 의한 난류혼합의 변화를 추정할 수 있다. 하지만 식생의 복잡한 형태에서 비롯한 흐름구조는 난류확산계수 변화에 미치는 영향 분석을 어렵게 한다.
식생으로 인한 흐름변화 및 혼합특성 간 관계를 분석하기 위해 실린더형 식생 모형을 이용한 많은 연구가 이뤄져 왔다. Nepf (1999)는 정수식생흐름 내 흐름방향으로의 확산을 기계적 확산(mechanical diffusion)과 난류확산(turbulent diffusion)의 합으로 정의했다. 이 연구에서는 기계적 확산이 식생으로 인한 불균일한 난류의 발생으로 인해 나타남을 보이며, 식생의 밀도를 이용하여 정의했다. 이후 White and Nepf (2003)은 기계적 확산을 식생 후면에서 발생하는 재순환류에 의한 혼합(vortex trapping)과 식생으로 인한 흐름 교란에 의해 발생하는 이차 후류에 의한 혼합(secondary wake)의 합으로 정의했다. 이러한 정의에 기초하여 횡 방향과 연직방향에 대한 난류확산계수의 결정 방안을 제시하였다. 실린더형 식생을 이용한 연구에서는 난류 특성길이가 실린더의 직경으로 대체하여 제시한 바 있었다. 하지만 Tanino and Nepf (2008)는 무작위적으로 식재 된 실린더형 식생 모형에 대한 실험을 통해 난류 특성길이를 식생 간 평균 간격과 식생의 직경의 상대적 크기 중 작은 값으로 정해야 함을 제시하였다. Yang (2024)은 식생흐름에 의한 난류확산계수 연구동향을 분석하며, 실린더형 식생에서 도출된 난류확산계수가 자연형 식생에서 도출된 결과와 상이함을 지적했다. 또한 Sonnenwald et al. (2017)은 같은 식생이라도 생장 시기에 따른 줄기 및 잎의 형태 변화가 난류확산계수에 변화를 줄 수 있음을 밝혀낸 바 있다. 또한 Xu and Nepf (2021)는 자연형 식생의 frontal area와 난류확산계수의 연직변화 간 상관관계를 분석하였으며, 이에 따라 난류특성길이를 식생의 frontal area의 변화에 따라 정의했다. 이러한 선행연구들은 자연형 식생의 형태가 흐름 구조와 난류 혼합 특성에 영향을 미친다는 점을 보여주었다. 그러나 자연형 식생은 성장 특성이 복잡하기 때문에, 식생을 구성하는 개별 요소가 흐름 및 혼합에 미치는 영향을 독립적으로 분석하는 데에는 한계가 있다.
침수식생 흐름에서는 자유흐름영역(flow zone)과 식생흐름영역(vegetation zone) 사이에서 발생하는 Kelvin-Helmholtz instability에 의해 coherent vortex structure가 발행하고, 이로 인해 발생하는 대규모 와류는 식생과 자유흐름영역 사이에서 물질교환을 촉진시키는 역할을 한다(White and Nepf, 2007; Nezu and Sanjou, 2008). 침수식생 상단에서 발생하는 전단흐름은 자유흐름과 식생 내부 흐름 간 운동량 차이에 의해 발생하며, 전단흐름의 강도는 식생의 특성(밀도, 유연성, 형태 등)에 따라 달라진다(Nepf, 2012). 이러한 식생 경계층에서 발생하는 전단흐름으로 인해 난류 운동에너지 및 Reynolds stress의 국부적 상승을 일으킨다(Luhar et al., 2008; Ghisalberti and Nepf, 2004). 침수식생에서 발생하는 흐름 구조의 변화는 난류 혼합 특성에도 영향을 미친다. 경계층에서 발생하는 전단흐름으로 인해 발생한 coherent vortex는 수직 방향의 난류확산을 증진시키며, 이는 오염물질, 부유사 등의 물질 이동에 영향을 미친다(Poggi et al., 2009; Tinoco et al., 2015). 침수식생 조건에서 발생하는 흐름구조 및 난류혼합 특성 변화 분석을 위해 많은 연구에서 실린더형 모형 식생을 이용한 연구를 수행해왔다(Baptist et al., 2007; Wong et al., 2020). 그리고 최근 연구에서는 자연형 식생을 이용하여 식생의 계절적 형태변화(가지 및 잎의 성장)가 흐름구조에 미치는 영향에 대한 연구가 수행된 바 있었다(Fonseca and Koehl, 2006; Vieira et al., 2023). 하지만 실제 자연형 식생은 가지, 잎, 줄기 등의 형태적 복잡성을 가지며, 이들 구성 요소가 난류 구조 및 혼합 특성에 미치는 영향을 구분하여 정량적으로 분석한 연구는 상대적으로 부족한 상황이다.
선행 연구를 통해 살펴본 바와 같이 식생의 형태적 특성(가지, 잎, 줄기 등)은 흐름 및 혼합 특성의 변화에 영향을 미칠 수 있음을 알 수 있었다. 따라서 본 연구에서는 식생의 형태적 특성 중 가지의 유, 무가 흐름 및 혼합특성 변화에 미치는 영향을 분석하기 위한 모형 실험을 수행했다. 이를 위해 가지가 없는 실린더형 식생 모형과 가지가 있는 식생 모형 내에서 유속의 시계열 변화를 측정하여 식생의 가지로 인해 발생하는 흐름특성을 분석했다. 그리고 시간 평균 된 유속의 연직분포와 난류흐름 특성을 비교하여 식생의 가지로 인한 흐름 특성 변화를 분석했다. 또한 흐름 측정결과로부터 난류확산계수를 산정하여, 식생의 가지가 식생 내부의 난류확산에 미치는 영향을 분석했다.
2. 연구방법
2.1 난류확산계수 산정
난류흐름 내에서 오염물질의 혼합거동은 3차원 이송-확산 방정식을 이용하여 계산할 수 있다.
여기서 는 시간평균 농도, 시간평균 유속, 는 난류확산계수이다. Eq. (1)의 난류확산계수는 이송-확산 방정식의 확산항을 조절하는 주요 계수임과 동시에 난류에 의한 오염물질의 혼합정도를 추정할 수 있는 지표이다. 난류확산계수는 Lagrangian statistics에 따른 입자 변위의 시간변화(Taylor, 1921) 또는 차원해석에 따른 난류확산계수는 유속 스케일과 난류특성길이의 곱(Nepf, 1999)으로 계산할 수 있다. Lagrangian statistics에 따라 오염물질 입자 간 거리() 제곱에 대한 ensemble mean ()은 입자 속도의 자기상관성(autocorrelation)을 잃을 만큼 충분한 시간이 흐른 뒤, 다음 식과 같이 유속 변동의 분산()과 integral time scale ()의 곱으로 나타낼 수 있다(Fischer et al., 1979).
여기서 이고 은 ensemble mean이다. Eq. (2)의 은 Ergodic 가정에 따라 고정된 지점에서 측정된 유속 변동의 시간평균()으로 대체할 수 있다. Integral time scale ()은 autocorrelation function ()으로부터 다음 식과 같이 계산한다.
여기서 는 autocorrelation function이 영점을 교차하는 시간이다. 이에 따라 Eqs. (2), (3), (4)로부터 난류흐름 특성으로부터 난류확산계수를 계산할 수 있다.
차원해석을 통한 난류확산계수의 계산은 다음 식과 같이 난류운동에너지의 제곱근()과 integral length scale ()를 이용하여 다음 식과 같이 계산할 수 있다(Nepf, 1999; Termini, 2019; Xu et al., 2022).
여기서 는 비등방성 난류 발생을 고려한 scale factor, 이다. Eq. (5)의 Integral length scale은 난류 흐름 내 대표적 와류의 평균적 크기를 의미하며, 다음 식과 같이 integral time scale과 시간평균 유속의 곱으로 계산된다(Mossa et al., 2017; De Serio et al., 2018; Brunet, 2020).
식생을 포함한 흐름에서 는 식생 주변에서 발생하는 난류의 스케일로 해석할 수 있다. 원형 실린더형 모형 식생의 경우 는 식생의 직경과 식생 간 거리 중 더 작은 값으로 정할 수 있다고 제안되었다(Tanino and Nepf, 2008). 반면, 자연형 식생에서는 가 식생의 형태적 특성에 따라 변화하는 값으로 나타난다고 알려진 바 있다(Xu and Nepf, 2020; Xu et al., 2022). 본 연구에서는 흐름 측정 결과로부터 Eq. (2)를 이용하여 난류확산계수를 산정하였으며, Eq. (5)의 관계로부터 무차원 난류확산계수()를 산정하여 식생 흐름에 의한 난류확산계수의 연직변화를 분석했다.
2.2 실험 방법
식생흐름 내 유속 측정을 위해 Fig. 1과 같은 길이 15 m, 폭 0.4 m의 사각 단면 개수로를 이용했다. 실험수로의 흐름은 유입부의 정류시설을 통과하여 안정화된 후 유입되며, 식생 영역은 안정화된 흐름조건을 확보하기 위해 유입부에서 6.5 m 떨어진 지점에 배치했다. 수로 내 식생 영역은 2 m 길이로 정하였으며, 스테인리스강(stainless steel)을 이용하여 제작한 모형식생을 이용하여 10 cm 간격으로 배치하였다. 이때 식생은 staggered 패턴으로 배열하여 식생 하류에서 흐름이 발달할 수 있는 공간을 확보하여 공간적으로 균일한 난류흐름이 발생하도록 했다. 식생 영역 내 배치된 식생은 130개이고 식생 식재 밀도는 162.5 m-2이다. 가지의 유, 무에 따른 흐름 변화를 비교하기 위해 모형에 사용한 식생은 직경() 3 mm, 높이() 0.1 m의 원형 실린더 형태(Type I)와 원형 실린더 식생 모형에 가지를 추가한 형태를 이용했다(Type II). 식생 수리실험을 위한 유량은 0.02 m3/s, 수심()은 0.2 m로 침수흐름조건을 나타내며, Fr 수는 0.21로 상류(subcritical flow) 조건을 나타낸다. 이 수심 조건에서 Type I과 Type II의 frontal area ()는 각각 0.244 m-1, 0.39 m-1이고, solid volume fraction (𝜙)은 각각 5.7×10-4, 9.2×10-4으로 가지 추가로 인해 Type II 조건에서 더 높은 식생영역을 차지함을 알 수 있다.
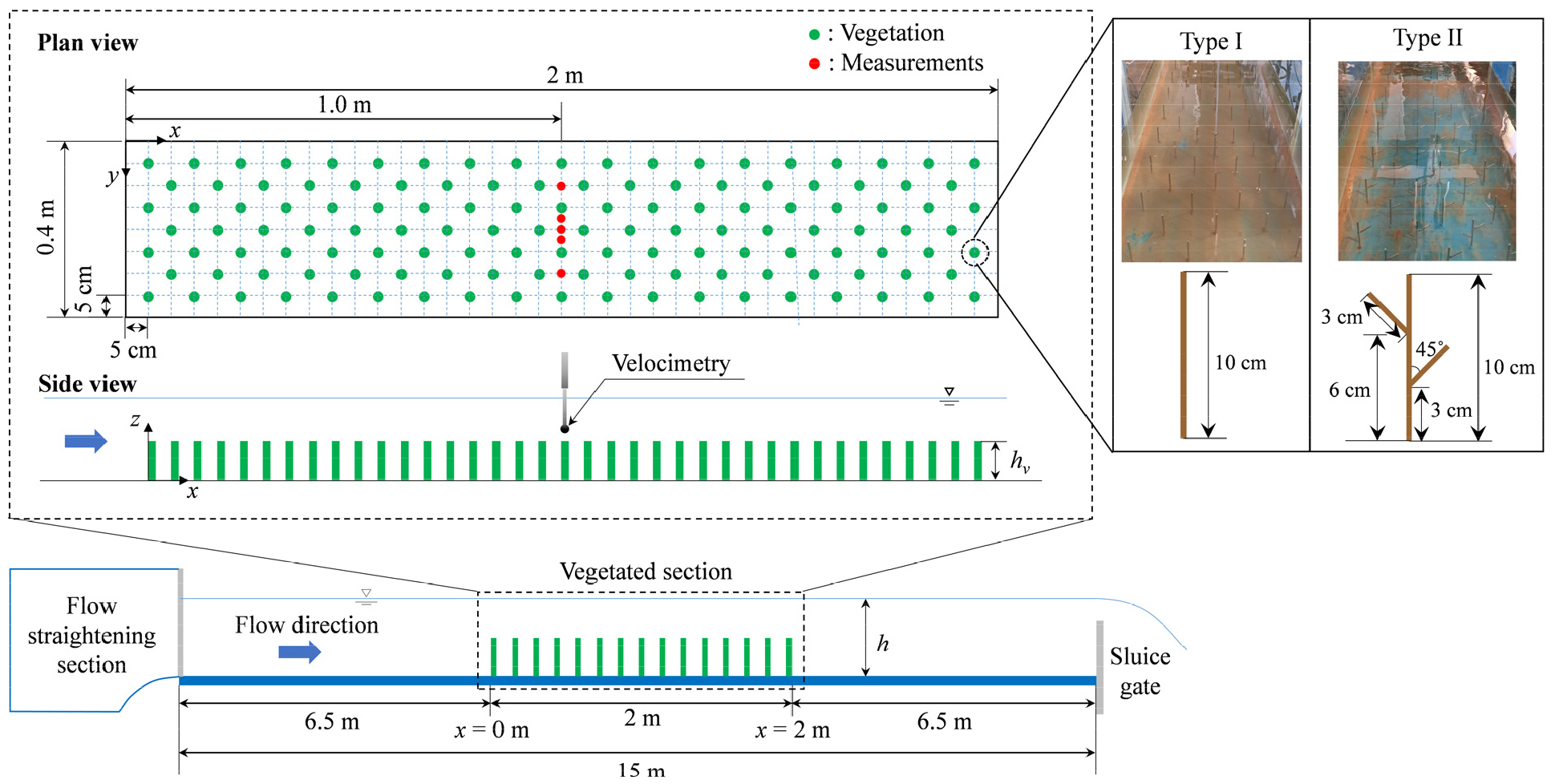
식생이 시작되는 지점에서는 급격한 흐름 변화에 따라 유속 및 수심 변화가 발생한다. 따라서 본 연구에서는 식생 구간의 상, 하류 0.6 m 범위 내 수심 변화가 1% 미만인 구간 내에서 유속측정 위치를 결정하였으며, Fig. 1과 같이 식생영역의 중앙(=2.5)에서 유속의 연직 변화를 측정했다. 그리고 하폭 방향으로 5개 지점(=0.25, 0.44, 0.50, 0.56, 0.75)에서 유속을 측정하였으며, 각 측점에서 수심방향으로 1 cm 간격으로 17개 지점에서 측정했다. 유속은 3차원 전자식 유속계(VP3000, KENEK)를 이용하여 흐름이 안정화된 후 측정했고, 각 측점 당 80Hz의 sampling rate로 60초 간 측정하여 x, y, z 방향 유속을 각각 4,800개씩 취득했다. 측정된 유속 시계열 데이터로부터 이상치 제거를 위해 유속의 시간 평균값으로부터 ±3𝜎를 벗어나는 값을 필터링한 뒤 분석에 활용했다.
3. 결과 및 토의
3.1 전단흐름 변화 특성
Fig. 2는 두 식생 유형에 대한 흐름방향 유속의 연직변화를 나타낸다. Fig. 2의 유속은 시간평균 유속()을 접근 유속()으로 나눈 값을 하폭 방향으로 평균()하여 나타냈다. 두 식생 유형 모두 식생으로 인한 항력 발생으로 유속이 감소하였으며, 접근흐름에 비해 식생 상단을 중심으로 급격한 유속의 연직변화를 나타냈다. 식생으로 인해 발생한 유속의 차이는 Kelvin-Helmholtz instability에 의한 와류를 일으키며, 운동량 전달을 유발하는 전단층을 형성한다. 이 전단층의 두께는 식생내부의 평균유속()과 자유흐름층의 평균유속()을 이용하여 정의할 수 있다. Ghisalberti and Nepf (2002)는 전단층의 두께를 =0.01에 해당하는 지점()으로부터 =0.01에 해당하는 지점() 간 거리로 정의하였으며, 여기서 이다. Table 1은 , , 의 계산 결과를 보여주며, 이 결과를 이용하여 계산한 Type I과 Type II의 전단층은 의 범위에서 발생했다. Type I의 경우 전단층()에서 유속이 완만하게 증가하는 경향을 보이는 반면, Type II에서는 전단층 내에서 Type I에 비해 급격한 유속 증가 경향을 보여준다. 이에 따라 유속경사()가 Type I에서 0.82 s-1인 것에 비해 Type II에서는 1.45 s-1로 더 급격한 유속 변화를 보여줬다.
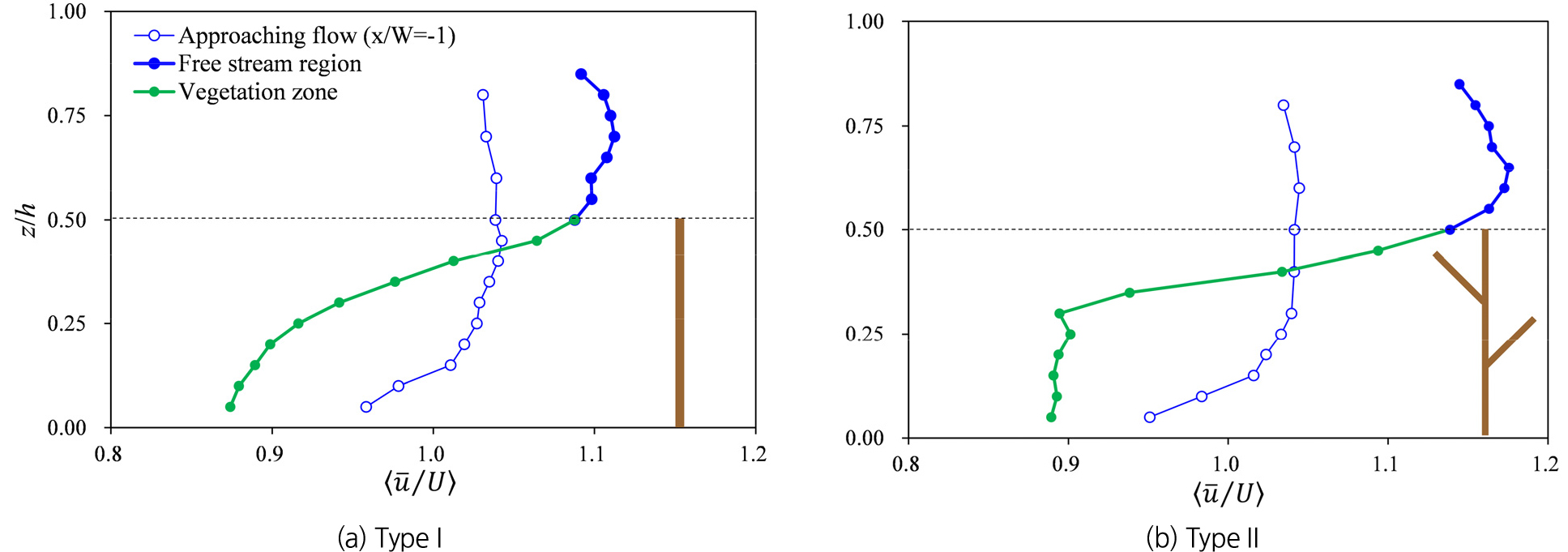
Table 1.
Velocity shear quantities according to vegetation types
| Vegetation | (m/s) | (m/s) | (m/s) | (m/s) | θ (mm) | λ |
| Type I | 0.284 | 0.246 | 0.038 | 0.265 | 5.660 | 0.072 |
| Type II | 0.312 | 0.256 | 0.055 | 0.284 | 3.107 | 0.098 |
Type I과 Type II 식생의 전단층 내 유속경사 차이는 식생 구조에 의해 형성된 운동량 분포(momentum distribution)의 차이와 관계되며, 이러한 운동량 분포 변화를 정량적으로 비교하기 위해 momentum thickness (𝜃)를 산정했다. 𝜃는 전단층에서 유속경사로 인해 발생한 운동량 변화를 등가 두께로 환산한 지표이다(Pope, 2000). 𝜃는 전단층 내() 유속변화로부터 다음 식과 같이 계산했다.
여기서 , , 는 와 의 평균이다. 그리고 전단층 상부와 하부의 유속차이로 인한 전단흐름의 상대적 세기(λ)는 로 정의된다(Caroppi et al., 2020). Type I과 II 식생의 𝜃와 λ 의 계산 결과는 Table 1과 같다. Type I과 II 식생의 전단흐름 특성을 비교한 결과, Type II에서 자유흐름층과 식생 내부 간 유속 차이()가 Type I보다 크게 나타났으며, 이로 인해 λ 또한 Type II 식생 흐름에서 더 강하게 형성됐다. 반면, θ는 Type II에서 더 얇게 계산되어, 전단흐름에 의한 운동량 변화가 Type I에 비해 좁은 영역에 집중되는 것으로 나타났다. 이러한 결과는 Type II 식생의 가지 구조가 식생 내부 유속을 Type I에 비해 더욱 감소시키고, 이에 따른 급격한 운동량의 연직변화가 전단층 내에서 강한 유속 경사를 유도한 것으로 해석할 수 있다. 이와 같이 가지에 의한 흐름구조의 변화는 전단층에서 난류흐름의 변화와 관계되며, 난류에 의한 혼합 양상 변화에 기여할 수 있다.
3.2 난류흐름 특성 변화
식생의 가지는 전단흐름 강화 및 운동량의 급격한 연직 변화를 일으킬 수 있으며, 이에 따라 난류 구조의 변화를 일으키게 된다. 따라서 식생 가지가 전단층 내 난류 구조에 미치는 영향을 분석하기 위해 난류운동에너지(), Reynolds 응력(), Shear production ()을 계산하여 Fig. 3에 나타냈다. Fig. 3에서 는 평균운동에너지(), Reynolds 응력은 , 는 난류에너지소산() 계산 결과와 각각 무차원화하여 비교했다. 이때 는 연직난류길이 스케일이며, Eq. (6)을 이용하여 계산했다. Type I과 II 모두 식생영역과 자유흐름영역의 경계에서 난류운동에너지()가 상승하는 결과가 나타났으며(Fig. 3(a)), Fig. 3(c)에서 볼 수 있듯 이 구간에서 가 상승하여 식생 상단의 전단흐름이 난류운동에너지 상승에 기여하고 있음을 알 수 있다. 또한 전단층 내 Reynolds stress의 증가는 경계층에서 연직방향으로의 운동량 교환이 활발히 이뤄지고 있음을 나타낸다. 하지만 θ에 대한 계산결과(Table 1)에서와 같이 Type II에서는 Reynolds stress의 상승이 Type I에 비해 좁은 영역에서 강하게 발생하고 있어, 운동량 교환이 식생 상단에 집중되어 발생하는 것을 볼 수 있다. 또한 Type II에서 난류운동에너지와 Reynolds stress가 Type I 보다 크게 나타났으며, 이는 식생의 가지에 의해 유발된 국지적 와류가 식생 내부에서 난류운동에너지의 상승과 운동량 교환을 활성화시키고 있음을 나타낸다. 반면, 난류에너지 소산 대비 shear production의 상대적 기여는 Type II보다 Type I에서 더 크게 나타났다(Fig. 3(c)). 이러한 결과는 Type I에서는 난류운동에너지가 주로 전단흐름에 의해 생성되지만, Type II에서는 shear production 외에도 식생 가지에 의해 발생한 소규모 와류 생성(wake production)이 전체 난류 운동에너지 증가에 기여하기 때문이다.
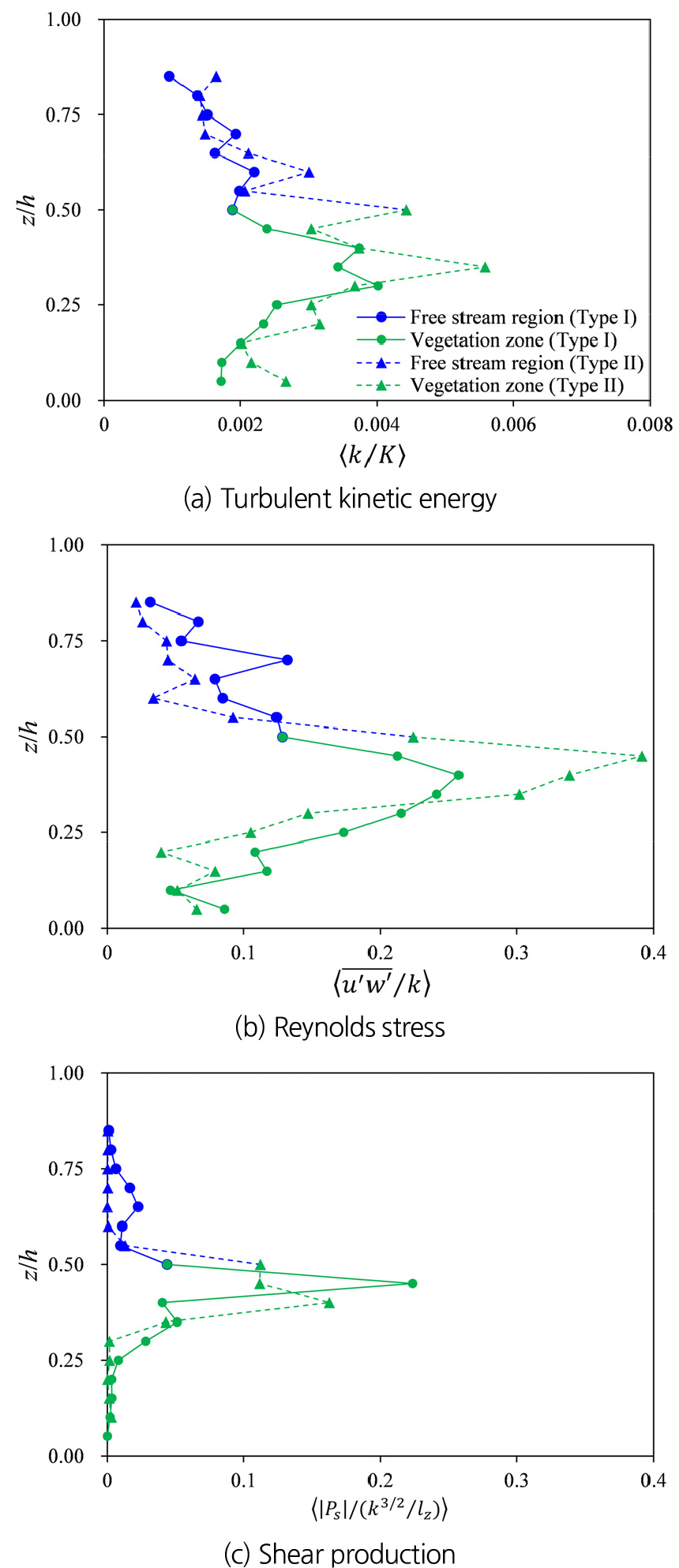
Fig. 4는 수로의 중심(y/W=0.5)에서 Eq. (4)를 이용하여 계산한 연직 속도 성분()에 대한 autocorrelation function ()의 시간변화를 보여준다. 두 식생 형태에서 모두 가 시간에 따라 감소하는 경향을 보였으며, 식생의 상부에서 가 진동하며 감쇠하는 패턴을 보였다. 이러한 의 진동ㆍ감쇠는 식생 상부 전단층에서 Kelvin-Helmholtz instability에 의해 생성된 와류가 주기적으로 생성, 소멸하며 coherent vortex structure가 유지되고 있음을 보여준다(Caroppi et al., 2019). Type II의 경우, Type I과 달리 식생 가지가 위치한 0.<<0.3구간과 자유흐름층 상부(0.8<<0.9)에서도 의 진동ㆍ감쇠 현상이 나타났다. 식생 내부에서 발생하는 진동ㆍ감쇠 현상은 가지에 의해 발생된 소규모 와류가 coherent vortex structure를 형성하여 발생한 것으로 해석된다. 반면, 자유흐름층 상부의 진동ㆍ감쇠는 전단층에서 생성된 coherent vortex structure가 자유흐름층으로 난류운동에너지를 상향 수송함으로써 유지된 것으로 볼 수 있다. Fig. 5에 나타낸 시간평균 된 난류운동에너지의 연직 flux ()계산 결과로부터 이러한 경향을 확인할 수 있다. Type II 식생은 Type I에 비해 식생 내부와 자유흐름영역 모두에서 연직 flux의 절대값이 크게 나타났으며, 특히 식생 상부에서 높은 상향 flux가 관찰되어, 0.8<<0.9구간의 coherent vortex structure 형성에 기여함을 알 수 있다.
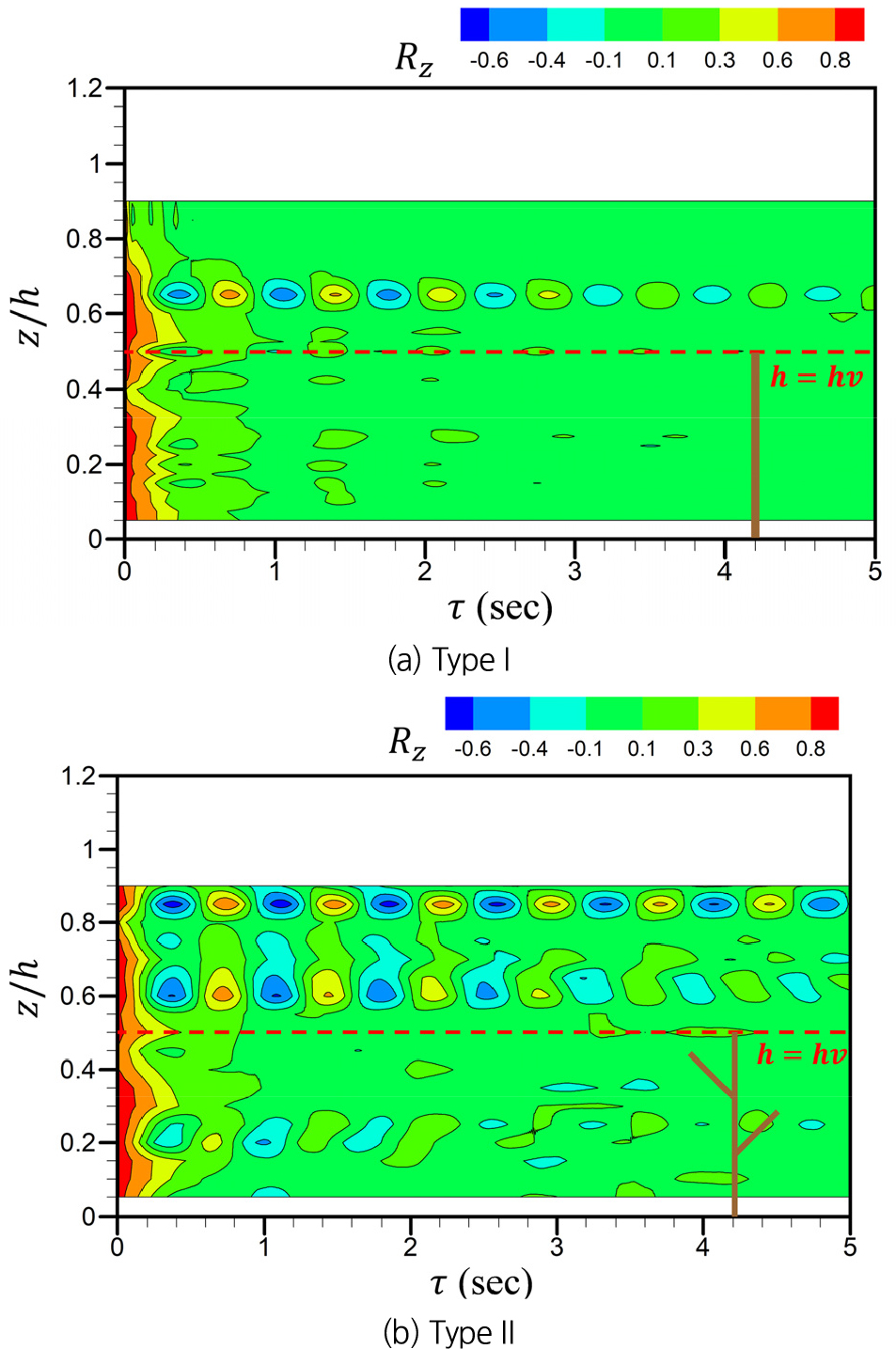
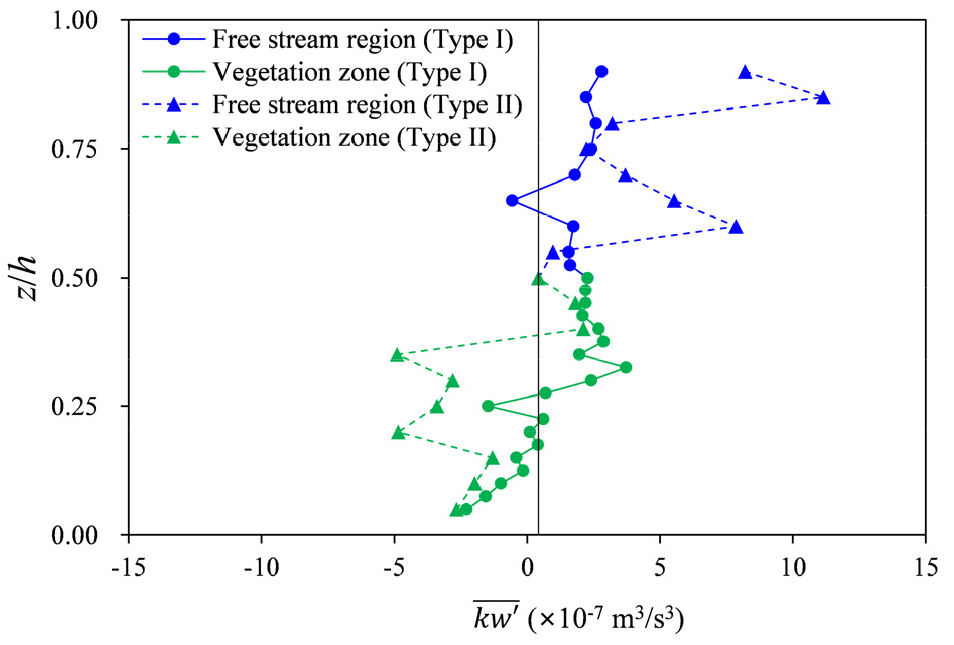
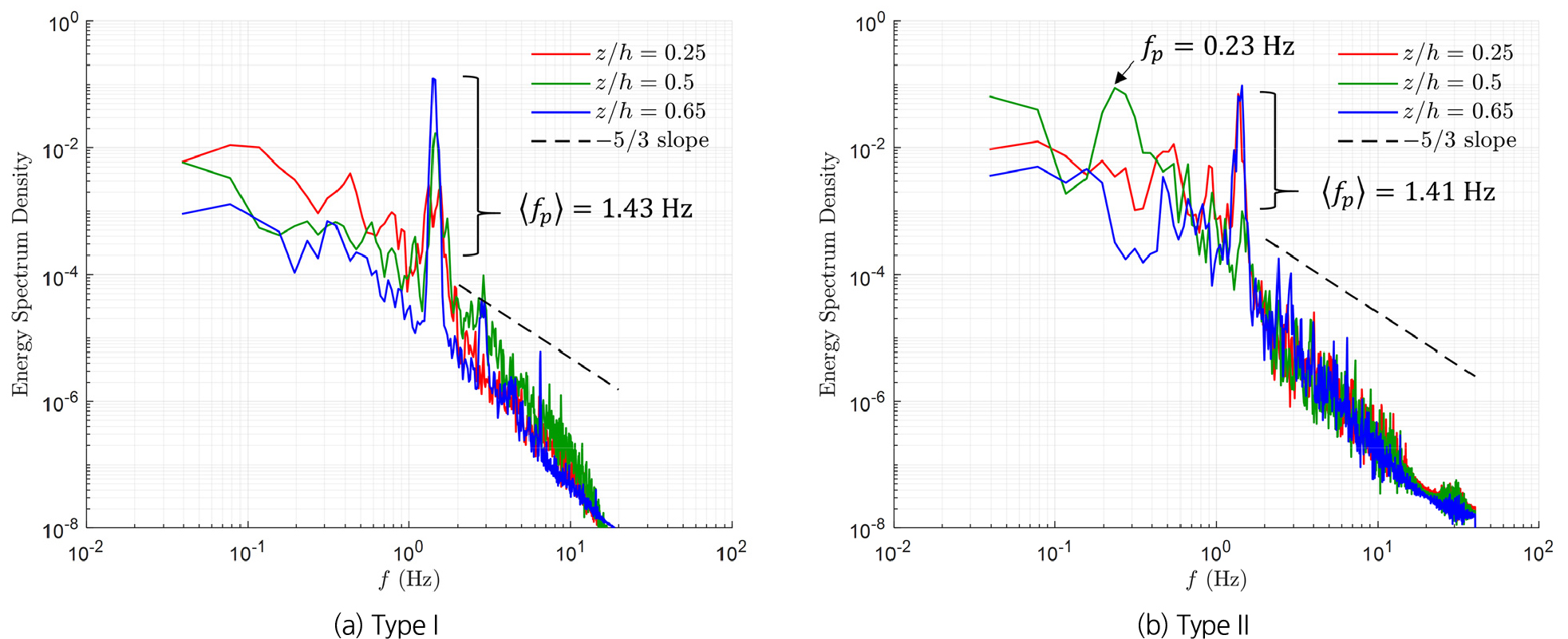
Fig. 6는 수로의 중심(=0.5)에서 를 이용하여 계산한 energy spectrum이며, 의 진동, 감쇠가 발생하는 {=0.25, 0.5, 0.65 위치에 대한 계산 결과를 보여준다. Energy spectrum으로부터 두 식생 유형 모두 특정 주파수 영역에서 에너지가 급격히 증가하는 경향을 보였고, 에너지가 최대값을 나타내는 주파수에 대한 평균 값()이 Type I에서는 1.43 Hz, Type II에서는 1.41 Hz로 유사하게 나타났다. 이러한 결과는 흐름이 식생을 통과하며 Fig. 4에서 확인한 바와 같이 coherent vortex structure를 형성하고 있음을 보여준다. 하지만 Type I 식생의 경우 식생영역과 흐름영역의 경계에 해당하는 =0.5에서도 첨두에너지가 관찰된 반면, Type II 식생의 경우 =0.5에서 첨두에너지가 상대적으로 낮은 주파수대()에서 나타났다. 이는 Type II 식생의 가지에 의해 범위에서 생성된 소규모 와류가 식생 상단의 coherent vortex를 작은 스케일의 와류로 분열시켜, 특정 주파수 대의 난류운동에너지 집중을 분산시킨 결과로 해석된다. 그 결과, 난류운동에너지가 더 작은 스케일의 난류로 전이되어 Type I에 비해 범위에서 난류운동에너지의 변동이 심화됐다. 이러한 고주파수대 에너지 변동 증가는 가지에 의해 형성된 소규모 와류가 지속적으로 작은 스케일 난류에 난류운동에너지를 공급하고 있음을 의미하며, 이는 Fig. 3(c)에서 추정한 바와 같이 식생의 가지로 인한 wake production 증가 경향과도 일치한다.
3.3 난류확산계수의 변화
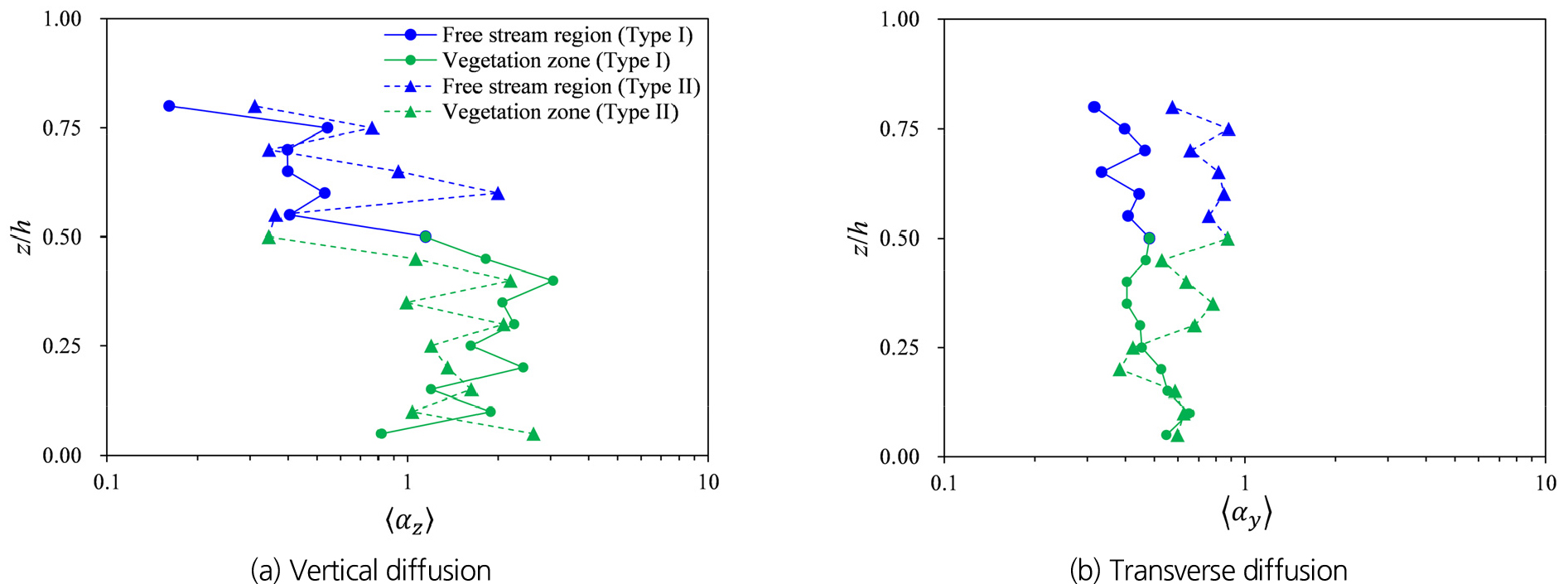
식생 가지로 인한 난류혼합의 연직변화 분석을 위해 Eq. (2)를 이용하여 횡 방향() 및 연직 방향 난류확산계수()를 산정했다. 그리고 Eq. (5)에 따라 계산된 난류확산계수를 으로 나눠 무차원화된 난류확산계수, 즉 scale constant()의 연직변화를 비교했다. Fig. 7은 Type I과 Type II 식생에 대해 무차원화된 난류확산계수()의 연직변화를 보여주며, 하폭방향으로 5개 지점에 대한 공간 평균 결과이다. Fig. 7(a)에서 연직 난류확산계수의 변화를 비교한 결과, 식생 영역에서는 Type I에 비해 Type II에서 가 감소하는 결과를 나타냈다. 이 결과는 가지로 인해 발생된 소규모 와류의 생성이 연직 난류확산을 Type I에 비해 감소시킨 것으로 해석할 수 있다. 특히 식생 상단(=0.5)에서는 coherent vortex structure가 식생 내 소규모 와류에 의한 분열로, 연직 혼합에 기여하는 난류 규모와 에너지가 감소하여 가 Type I의 약 30% 수준으로 감소하였다. 반면, 횡 방향 난류확산계수()는 =0.25~0.5 범위에서 Type I보다 더 높은 난류확산을 나타냈다. 이 결과로부터 식생 가지로 인해 연직 방향에 대한 난류확산의 감소 및 횡 방향 난류확산이 증가함을 알 수 있다. 자유흐름영역(>0.5)에서는 횡 방향과 연직 방향 모두 Type I보다 Type II 식생에 대해 더 큰 난류확산을 나타냈다. 이 결과는 Fig. 5에서 확인한 바와 같이 Type II 식생에서 생성된 난류운동에너지가 높은 상향 flux를 통해 자유흐름영역으로 더 활발히 전달되어, 난류확산을 유발하는 난류 규모의 발달에 기여했기 때문이다.
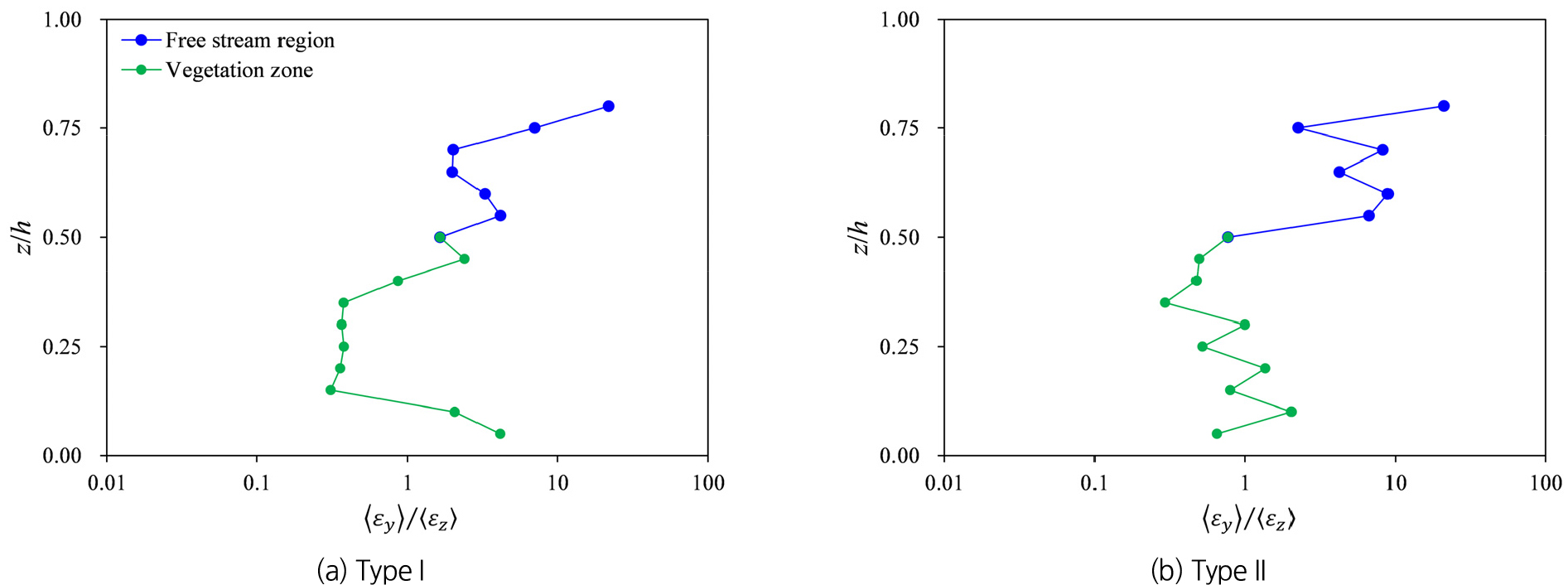
Fig. 7에서 살펴본 식생 가지로 인한 난류확산의 차이는 오염물질 또는 부유사의 혼합 및 체류시간에 영향을 미칠 수 있다. 가지가 발달한 식생 흐름에서는 식생 상단에서 연직 난류확산이 감소하므로 오염물질이나 부유사의 식생 내부로의 침투가 어려워진다. 반면, 이미 식생 내로 유입된 오염물질이나 부유사는 가지에 의해 연직 방향의 혼합이 억제되어 식생 내부에 상대적으로 더 오랜 기간 체류할 가능성이 높아진다. 따라서 식생이 하폭 전반에 걸쳐 분포되어 있는 경우, 하폭 방향보다는 연직 난류확산이 오염물질 배출에 더 큰 역할을 수행하게 되며, 가지의 존재로 인해 식생 내부에서 식생 외부로의 연직 방향 배출이 제한될 수 있다. 이러한 식생 가지에 의한 횡 방향과 연직 방향 혼합의 상대적 차이를 정량적으로 평가하기 위해, Fig. 8에서는 횡 방향과 연직 방향 난류확산의 비()를 비교했다. 그 결과, 두 식생 유형 모두 자유흐름 영역에서는 가 에 비해 크게 계산된 반면, 식생영역에서는 대체로 가 보다 작게 계산되었다. Type I의 경우 0.15≤<0.4 영역에서 ≈0.36으로 나타나 연직 난류확산이 일관되게 더 큰 결과를 보였다. Type II에서는 동일영역에서 ≈0.74로 계산되어 연직 난류확산이 더 우세하지만 일부 구간에서 횡 방향 난류확산이 더 크게 발생할 수 있음을 보여준다. 따라서 식생 가지로 인해 식생 내부에 체류하는 오염물질 또는 부유사가 횡 방향으로 더 활발하게 확산될 수 있다는 것을 보여준다.
4. 결 론
본 연구에서는 침수식생 흐름 조건에서 식생의 가지가 난류흐름 구조에 미치는 영향을 분석했다. 가지가 없는 식생 유형(Type I)과 가지가 있는 식생 유형(Type II)에 대해 유속의 연직 변화 측정 결과로부터 시간 평균 유속과 난류흐름 특성을 분석하여 가지에 의한 흐름구조의 변화를 비교했다. 실험결과로부터 도출된 주요 결과는 다음과 같다.
(1)Type II 식생에서는 Type I 식생에 비해 식생 내부에서 급격한 유속 변화가 발생하여 상대적 전단흐름 세기(𝜆)가 더 크게 나타났고, 식생과 자유흐름영역의 경계에서 momentum thickness가 Type I 보다 얇게 형성되어 운동량 변화가 좁은 영역에 집중되었다. 이 결과는 식생의 가지가 식생 내부 유속을 강하게 감속시키며, 경계에서 집중적인 전단층을 형성함을 보여준다.
(2)Type II에서는 가지에 의한 소규모 와류 발생으로 인해 Reynolds 응력 및 난류운동에너지가 Type I보다 크게 나타났다. 이 결과로부터 식생 내부에서 가지에 의한 난류 생성 메커니즘이 전단흐름뿐 아니라 난류흐름 구조 차이 발생에 기여함을 확인할 수 있다.
(3)Autocorrelation과 energy spectrum 분석 결과, Type I 식생에서 발생되는 coherent vortex structure가 Type II 식생에서는 가지에 의해 분열될 수 있음을 보여줬으며, 이에 따라 식생 상단에서 발생하는 Kelvin-Helmholtz vortex의 발생을 억제할 수 있음을 보여줬다.
(4)Type II 식생에서는 연직 방향 난류확산이 Type I에 비해 식생 상단에서 약 70% 수준까지 감소하였으며, 이는 가지 구조에 의해 발생한 소규모 와류가 연직 혼합을 억제한 결과이다.
(5)Type II 식생에서는 Type I에 비해 가 식생 내부에서 큰 값을 나타내어 식생 내 오염물질 또는 부유사의 연직 혼합보다 횡 방향 혼합이 더 활발하게 발생될 수 있음을 나타냈다.
이러한 결과로부터 식생의 가지 구조가 난류흐름구조와 함께 난류 혼합 특성 변화에 영향을 미칠 수 있음을 알 수 있다. 하지만 이 연구에서 적용한 가지의 형태는 제한적이며, 가지의 발생이 흐름 구조에 미치는 영향을 정량화하기 위해서는 다양한 형태의 가지 구조를 모사한 추가적 실험이 필요하다. 또한 자연형 식생의 잎과 줄기의 연성 특성은 주기적 흐름 변화를 유발하여 난류 구조에 영향을 미칠 수 있으므로(Caroppi and Järvelä, 2022), 가지 발달과 함께 잎의 유무 및 연성 조건에 따른 난류 혼합 변화에 대한 추가적 연구도 필요하다. 이와 같이 식생을 포함한 자연하천에서 오염물질 거동 예측 정확도 향상을 위해 식생의 형태적 특성을 반영한 흐름 및 혼합특성 변화에 대한 검토가 필요하다.
